【骨骼肌细胞膜与疲劳】
拼译:fatigue and cellular memlsane of skeletal muscle
20世纪80年代一些学者,如埃得瓦尔德(Edwazds)于1983年,坚耐斯(Jones)于1986年,柯泼尔(Coppen)和泰必特(Tibbits)于1988年相继指出骨骼肌细胞膜的机能变化常与运动性疲劳的发生有密切关系,诱发疲劳的机制几乎都是因为兴奋性衰竭或兴奋诱导的收缩机能的衰竭,其变化特点与刺激的频率(运动强度)有关。
肌细胞膜对细胞内环境的恒定、能量的转换、刺激的传递、细胞的生存和机能、与细胞外环境作用等都起着重要的作用。肌细胞膜通过几种途径调节代谢,依靠膜上的物质转运器转运葡萄糖、脂肪酸等能源物质。一些膜蛋白,如胰岛素和肾上腺素能受体可以产生第2信使环腺苷酸(CAMP),三磷酸肌醇(IP3)和钙离子(Ca2+),调节物质转运器的机能和肌细胞代谢,但至今尚未证明在疲劳的发生过程中这些转运器的机能不足或紊乱所起的重要作用。肌细胞正常的H+浓度约100nmol/L,对保持其正常生理机能极其重要,1983年道森(Dawson)指出在激烈运动时肌细胞H+浓度显著下降,最大可以升至1000nmol/L,这种重度的酸中毒被许多学者认为是导致疲劳的重要原因,尽管曾提出多种设想,但至今对酸中毒如何诱发疲劳的机制了解得并不透彻。运动时引起肌细胞酸中毒最常见的原因是糖无氧酵解产生过量乳酸。虽然骨骼肌也是人体内调节H+浓度的重要部位,但这一机能常被忽视。静息状态的骨骼肌,氢离子(H+)经过细胞膜也并非被动的电化学平衡,例如细胞外液H+浓度为10-7.4,细胞内液为10-7.0,经计算H+的平衡电位约为-23mV。静息状态骨骼肌细胞膜极化状态膜电位约-80mV。因此,有极强的电化学势能使H+从细胞外液流入胞内,H+必需经过主动转运机制运到胞外维持胞内H+浓度。H+运出肌细胞通过Na+/H+反向转运器,受到膜钠电位(ENa)和氢电位(EH)的调节,肌细胞内H+浓度高刺激Na+进入细胞。非特异的阴离子反向转运器(或Cl-/HCO3-转运器)对逆转细胞内酸化、调节骨骼肌细胞内H+浓度的生理意义不大。在进行激烈运动时最具有重要作用的是乳酸转运器,其作用是调节乳酸进出骨骼肌及心肌等细胞。乳酸的PKa为3.9,在生理H+浓度99%以上的乳酸都电解成为H+和乳酸根阴离子(La-),这种荷电的离子不易通过细胞膜,肌细胞膜上有乳酸转运器进行易化转运,它对乳酸的km约为20mmol,表现为La-/H+同向转运,其转运动力学包括方向是受限制的,La-和H+顺浓度梯度越过细胞膜,并产生饱和动力学效应。激烈运动时肌细胞内积聚的H+和La-均直接由电化学势能调控,固向转运出肌细胞,延缓疲劳的发生。激烈运动时或运动训练对肌细胞膜乳酸转运器产生何种影响至今还一无所知。激烈运动时肌细胞H+浓度显著升高,发生酸中毒,可以认为是H+的生成速率超过了H+跨膜转运的速率,与疲劳的发生有密切的关系。和其他类型的兴奋组织相似,骨骼肌至少有3种膜蛋白调节钙离子(Ca2+)的跨膜转运:电压-依赖Ca2+通道,Ca2+-ATP酶,Na/Ca2+交换器。在生理条件下后两种蛋白的作用似乎只使Ca2+流出肌细胞外,而Ca2+通道的作用只使Ca2+流入胞内。虽然,现在对骨骼肌收缩时细胞外液Ca2+跨膜流入胞内已很少还有疑问,但对它在兴奋收缩时的偶联作用仍有争议。有人认为肌肉收缩的力量可受细胞外液不同浓度的Ca2+影响,胞外Ca2+进入肌细胞内对诱发肌质网释放Ca2+是重要的,肌肉收缩时Ca2+进入肌纤维速度为每分钟每根肌纤维1.5PM。但另一些学者认为肌细胞外Ca2+进入肌纤维对激发收缩无作用,理由是虽然Ca2+流入肌细胞与收缩有联系,但发生于K±诱导收缩后,Ca2+流入量极小,每英尺肌质网每秒小于1个Ca2+,不可能诱发收缩。肌细胞膜除极化后膜电位超过Ca2+平衡电位,又引起Ca2+流向胞外,因此,胞内Ca2+的瞬时值大体未变,肌细胞仍然收缩。现有人认为肌细胞外Ca2+流入可补充胞内钙库和激活代谢,尤其是激性磷酸化酶加速糖酸解。横管的二羟吡啶(DHP)受体的密度高于其他组织10倍,其中一小部分为机能性Ca2+通道,大部分作为横管的电压感受器,某种调节物质如IP3可能在横管电压感受器与肌质网释放Ca2+起信使作用。已证明快肌纤维横管DHP受体及Ca2+流入量都比慢肌纤维多。骨骼肌Ca2+通道也可受儿茶酚胺经CAMP调节引起钙电流增加和正变力反应。骨骼肌细胞机能紊乱是导致疲劳的重要原因,出现细胞内液K+浓度降低,Na+、CL-浓度升高,细胞外液K+浓度升高,pH降低细胞膜电位下降,以及肌细胞兴奋性和收缩性的变化等。肌细胞内液K+浓度比细胞外液高30倍左右,细胞外液Na+浓度比细胞内液高14.2倍,主要依靠膜钠泵不断工作来维持静息膜外内外浓度梯度,从理论上说每个钠泵每分钟可以泵入膜内16000个K+,运动时血浆儿茶酚胺浓度升高,通过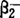 肾上腺素受体激活钠泵,使之活性比静息时增高3倍,但往往还不能满足大强度运动时的需要,在疲劳过程中钠泵的机能受到明显抑制,使肌细胞丢失K+比获得Na+要多3倍。近年来核磁共振研究表明接近细胞膜的游离的ATP浓度下降就可以增大K+传导性,加速细胞内K+进入细胞外液。因此,大强度运动引起疲劳的表现为膜电位降低,动作电位构象改变,肌质网释放Ca2+减少,最终导致肌肉收缩力量下降。但是这种解释疲劳的机制也有一些争议,因为在运动后的恢复期中细胞内K+浓度、膜电位和动作电位构象的恢复比肌肉收缩力量的恢复速度要快得多。Ca2+浓度的变化也和疲劳的发生有关,大强度运动会使Ca2+在横管中积聚,使兴奋阈提高易产生横管传播动作电位的机能衰竭,并可能产生Ca2+从肌质网中移位,肌质网中Ca2+排空。长期以来人们认为大强度运动时骨骼肌细胞内H+浓度升高是导致肌肉收缩时力量降低的重要因素,运动后肌肉收缩力量的恢复与H+浓度的恢复呈高度相关性。近年来研究用电刺激蛙缝匠肌产生疲劳,发现肌肉收缩力量下降虽然不受细胞外液液中不同H+浓度的影响(H+浓度10~398.1nmol/L),但溶液中H+浓度降低可以使肌肉张力恢复速度加快,膜电位、动作电位构象的恢复也加快,乳酸流出肌肉的速度加快,但如果用桂皮酸盐阻断La-/H+同向转器就会使疲劳肌中乳酸流出的速度减慢。高强度电刺激引起疲劳的肌肉中H+浓度恢复速度比K+或Na+要慢。在运动实践的研究中经常报导服用碳酸氢钠等碱性药物并不能提高大强度运动的工作能力,但对短时间内重复进行第2次大强度运动的能力有所改善,可能与上述研究结果吻合。现已证明运动性疲劳具有兴奋收缩过程的解偶联作用,正常的兴奋不能引起肌质网Ca2+释放和肌纤维收缩。由于横管的兴奋与收缩偶联的机制至今还未完全明了,也难以正确推测解偶联的机制,根据目前DHP受体的大部分可作为电压感器诱导IP3的释放和进而引肌质网终池Ca2+通道开放的假定来分析,在疲劳过程中IP3的调节也可能产生紊乱,影响了DHP敏感Ca2+通道的机能。近来的研究还发现PH降低可以使DHP敏感Ca2+通道质子化。使得这种单通道对Ca2+的传导性降低3倍,产生了肌质网释放过程的部分阻断作用。至于疲劳过程中如何影响DHP电压感受器,电压感受器在兴奋收缩偶联中的作用仍有待于进一步研究。虽然近年来一些研究发现了高强度或低强度运动中肌细胞膜的某些机能变化,但目前还很难确定这些变化在肌肉疲劳中所占地位与因果关系。1988年泰必特(Tibbit)提出除了上面所讨论的诸因素外,低强度运动所产生疲劳可能还和磷脂酶激活和自由基生成增多诱发脂质过氧化反应及蛋白质交联影响了膜结构完整性和转运系统的机能等有关。近年来肌细胞膜在疲劳发生过程中的作用逐渐引起了人们的关注,并发始逐渐深入地进行研究。【参考文献】:1 Gneen H J.Human Kinetics Books Champaign.Iiinois,1988,13~352 Tibbits G F.Human Kinetics Books Champaign.Iiinois,1988,37~573 许豪文.运动生物化学进展.上海:华东师范大学出版社,1990.146~1554 Housh J J,et al.Int.J.Sponts Med,1991,3(12):305~308
肾上腺素受体激活钠泵,使之活性比静息时增高3倍,但往往还不能满足大强度运动时的需要,在疲劳过程中钠泵的机能受到明显抑制,使肌细胞丢失K+比获得Na+要多3倍。近年来核磁共振研究表明接近细胞膜的游离的ATP浓度下降就可以增大K+传导性,加速细胞内K+进入细胞外液。因此,大强度运动引起疲劳的表现为膜电位降低,动作电位构象改变,肌质网释放Ca2+减少,最终导致肌肉收缩力量下降。但是这种解释疲劳的机制也有一些争议,因为在运动后的恢复期中细胞内K+浓度、膜电位和动作电位构象的恢复比肌肉收缩力量的恢复速度要快得多。Ca2+浓度的变化也和疲劳的发生有关,大强度运动会使Ca2+在横管中积聚,使兴奋阈提高易产生横管传播动作电位的机能衰竭,并可能产生Ca2+从肌质网中移位,肌质网中Ca2+排空。长期以来人们认为大强度运动时骨骼肌细胞内H+浓度升高是导致肌肉收缩时力量降低的重要因素,运动后肌肉收缩力量的恢复与H+浓度的恢复呈高度相关性。近年来研究用电刺激蛙缝匠肌产生疲劳,发现肌肉收缩力量下降虽然不受细胞外液液中不同H+浓度的影响(H+浓度10~398.1nmol/L),但溶液中H+浓度降低可以使肌肉张力恢复速度加快,膜电位、动作电位构象的恢复也加快,乳酸流出肌肉的速度加快,但如果用桂皮酸盐阻断La-/H+同向转器就会使疲劳肌中乳酸流出的速度减慢。高强度电刺激引起疲劳的肌肉中H+浓度恢复速度比K+或Na+要慢。在运动实践的研究中经常报导服用碳酸氢钠等碱性药物并不能提高大强度运动的工作能力,但对短时间内重复进行第2次大强度运动的能力有所改善,可能与上述研究结果吻合。现已证明运动性疲劳具有兴奋收缩过程的解偶联作用,正常的兴奋不能引起肌质网Ca2+释放和肌纤维收缩。由于横管的兴奋与收缩偶联的机制至今还未完全明了,也难以正确推测解偶联的机制,根据目前DHP受体的大部分可作为电压感器诱导IP3的释放和进而引肌质网终池Ca2+通道开放的假定来分析,在疲劳过程中IP3的调节也可能产生紊乱,影响了DHP敏感Ca2+通道的机能。近来的研究还发现PH降低可以使DHP敏感Ca2+通道质子化。使得这种单通道对Ca2+的传导性降低3倍,产生了肌质网释放过程的部分阻断作用。至于疲劳过程中如何影响DHP电压感受器,电压感受器在兴奋收缩偶联中的作用仍有待于进一步研究。虽然近年来一些研究发现了高强度或低强度运动中肌细胞膜的某些机能变化,但目前还很难确定这些变化在肌肉疲劳中所占地位与因果关系。1988年泰必特(Tibbit)提出除了上面所讨论的诸因素外,低强度运动所产生疲劳可能还和磷脂酶激活和自由基生成增多诱发脂质过氧化反应及蛋白质交联影响了膜结构完整性和转运系统的机能等有关。近年来肌细胞膜在疲劳发生过程中的作用逐渐引起了人们的关注,并发始逐渐深入地进行研究。【参考文献】:1 Gneen H J.Human Kinetics Books Champaign.Iiinois,1988,13~352 Tibbits G F.Human Kinetics Books Champaign.Iiinois,1988,37~573 许豪文.运动生物化学进展.上海:华东师范大学出版社,1990.146~1554 Housh J J,et al.Int.J.Sponts Med,1991,3(12):305~308(华东师范大学许豪文教授撰)