【ATP酶的结构与功能】
拼译:structure and function of ATPase
ATP是生物体内各种生命活动的直接能源和间接的能源,一个成年人在静息状态时,每天消耗约70kgATP,在进行重体力劳动时,每天消耗约800kg以上。但是,ATP不能透过细胞质膜,在细胞内通过ATPase催化ATP分解与合成的偶联、循环,使ATP保持恒量。生物体需要的能量愈多,循环的速度就愈快,ATPase活性直接影响循环的速度。多年研究资料证明,ATPase的种类很多,而且所有的ATPase都是与生物膜(细胞质膜和细胞器膜)相结合,与离子的跨膜转运有关。现在知道的至少有三类离子转运ATPase。
P-ATPase:这类ATPase的特点是酶在反应过程中形成共价磷酸化的中间物,由于钒酸盐可以代替磷酸与酶共价结合,所以钒酸盐是这类酶的专一性抑制剂。P-ATPase分布在真核细胞的质膜、少数细胞器膜和大肠杆菌、链球菌的内膜。大多数PATPase只有一条具有催化活性约70000~100000,另一条催化活性为55000。真核生物(酵母、真菌如脉胞杆菌和动植物)的质膜ATPase催化ATP水解的同时,泵质子(H+)到细胞外形成质子电化学梯度(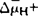 ),产生质子驱动力(Proton Motive Force),简称PMF(
),产生质子驱动力(Proton Motive Force),简称PMF(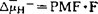 ,F:法拉第常数,
,F:法拉第常数,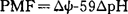 ,ψ:膜电位),为糖、氨基酸和阴离子、阳离子的主动跨膜转运提供动力,称为质子泵ATPase(H+-APTase)。阳离子激活该类酶活性。对多种P-ATPase考察证明,在具有催化活性的多肽中,有8个疏水段横跨质膜磷脂双分子层,各段之间有亲水肽段连接,多肽的N、C末端和大部分的亲水肽段位于质膜的内侧,每两个疏水肽段组成一个区,分别称为I、Ⅱ、Ⅲ、Ⅳ区,第Ⅱ和第Ⅲ区相距较远,两者之间的亲水肽段最长,含有可发生磷酸化的部位和几种抑制剂如NEM(N乙基顺丁烯二酰亚胺,专一性修饰疏基的试剂)等的结合位点,过量的ATP可保护这些位点不与抑制剂结合。质子通道的抑制剂DCCD(二环已基碳二亚胺)结合位点是第Ⅰ区Giu129。P-ATPase在反应过程中,经过E1和E2两种构象变化,发生磷酸化作用水解ATP,为此,常把P-ATPase称为E1E2类型ATPase。一级结构、基因克隆和序列测定工作进一步证明,各种P-ATPase有着类似的结构和作用机制,它们之间的差异在于转运不同的离子,Na+K+-ATPase使Na+、K+逆向跨膜交换,其它的酶则分别转运Ca2+、H+和K+,这是因为在这些酶分子中,与离子结合或形成离子通道部位的氨基酸种类不同,因而它们对于抑制剂的敏感性也不同,抑制Na+K+-ATPase的鸟本苷不抑制H+,Ca2+ATPase。F-ATPase:这是一类结构最复杂、研究最深入的质子泵ATPase,存在于细菌内膜、叶绿体的类囊体和线粒体内膜的内侧,分子量450kda以上,由水溶性的F1、脂溶性F0和连接F1与F0的柄状寡霉素敏感蛋白(OSCP:Oligomycin SensitivityConferring Protein)3部分组成。不同来源的F-ATPase含有数量不等的亚基:细菌8个,叶绿体8~9个,酵母和鼠肝10个,牛心13个。早在1960年,美国康奈尔大学的RakerE.,教授及其同事用机械搅拌的方法使牛心线粒体ATPase的可溶性部分脱落下来,进行纯化和重组研究,定名为F1。电镜观察到F1组分为直径9nm的椭圆形球体,分子量360000~380000。F1组分由α、β、
,ψ:膜电位),为糖、氨基酸和阴离子、阳离子的主动跨膜转运提供动力,称为质子泵ATPase(H+-APTase)。阳离子激活该类酶活性。对多种P-ATPase考察证明,在具有催化活性的多肽中,有8个疏水段横跨质膜磷脂双分子层,各段之间有亲水肽段连接,多肽的N、C末端和大部分的亲水肽段位于质膜的内侧,每两个疏水肽段组成一个区,分别称为I、Ⅱ、Ⅲ、Ⅳ区,第Ⅱ和第Ⅲ区相距较远,两者之间的亲水肽段最长,含有可发生磷酸化的部位和几种抑制剂如NEM(N乙基顺丁烯二酰亚胺,专一性修饰疏基的试剂)等的结合位点,过量的ATP可保护这些位点不与抑制剂结合。质子通道的抑制剂DCCD(二环已基碳二亚胺)结合位点是第Ⅰ区Giu129。P-ATPase在反应过程中,经过E1和E2两种构象变化,发生磷酸化作用水解ATP,为此,常把P-ATPase称为E1E2类型ATPase。一级结构、基因克隆和序列测定工作进一步证明,各种P-ATPase有着类似的结构和作用机制,它们之间的差异在于转运不同的离子,Na+K+-ATPase使Na+、K+逆向跨膜交换,其它的酶则分别转运Ca2+、H+和K+,这是因为在这些酶分子中,与离子结合或形成离子通道部位的氨基酸种类不同,因而它们对于抑制剂的敏感性也不同,抑制Na+K+-ATPase的鸟本苷不抑制H+,Ca2+ATPase。F-ATPase:这是一类结构最复杂、研究最深入的质子泵ATPase,存在于细菌内膜、叶绿体的类囊体和线粒体内膜的内侧,分子量450kda以上,由水溶性的F1、脂溶性F0和连接F1与F0的柄状寡霉素敏感蛋白(OSCP:Oligomycin SensitivityConferring Protein)3部分组成。不同来源的F-ATPase含有数量不等的亚基:细菌8个,叶绿体8~9个,酵母和鼠肝10个,牛心13个。早在1960年,美国康奈尔大学的RakerE.,教授及其同事用机械搅拌的方法使牛心线粒体ATPase的可溶性部分脱落下来,进行纯化和重组研究,定名为F1。电镜观察到F1组分为直径9nm的椭圆形球体,分子量360000~380000。F1组分由α、β、 、δ和ε5种亚基组成(
、δ和ε5种亚基组成(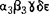 ),不同来源的F-ATPase复合物中,F1组分差别很小,α亚基含有合成ATP的活性部位,
),不同来源的F-ATPase复合物中,F1组分差别很小,α亚基含有合成ATP的活性部位,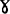 亚基含有分解ATP的活性部位,β亚基含有ATP的结合位点,此外还有一条称之为ATPase抑制因子的多肽,与F1可逆性的结合。F0组分是与膜结合的脂蛋白,不同来源的F-ATPase复合物中,F0组分的亚基数差别较大,E.Coli的F0有a.b.c三种亚基,c为DCCD结合亚基,每个F0中至少有10个c亚基,和a、b亚基结合形成质子通道。真核生物的Fo通常含有5~8个亚基。全酶具有催化ATP合成与分解双重活性,受寡霉素、叠氮化钠和DCCD的抑制。当ATPase抑制因子与F1结合时,该酶催化ATP合成,反之,催化ATP水解。除机械搅拌外,用EDTA、氯仿处理,也可使F1组分脱落下来,可溶性F1保留催化ATP水解的活性,被称为溶性F1-ATPase,对寡霉素和DCCD不敏感,低温下迅速失活(Coldlabile),其它性质与全酶相似。各种F-ATPase的差异在对抑制剂的敏感性不同。细菌的H+-ATPase对寡霉素不敏感,连接F1和F0的不是OSCP,而是被称之为连接素或结合素(nectin)的蛋白质。叶绿体H+ATPase对寡霉素的敏感性不如线粒体H+对ATPase强,有些甚至完全不敏感。V-ATPase:这类ATPase因存在于真核细胞内的空泡型(Vacuolar)细胞器上而得名,如酵母、真菌的空泡,植物的液泡,动物的溶酶体、笼蛋白衣被小泡(Clathrin Coated Vesicles)、嗜铬颗粒(贮存肾上腺素等激素)、突触小泡(贮存乙酰胆碱)和分泌小泡等。V-ATPase是3类离子转运ATPase中数量最多的一类,它们虽有许多共性,但在它们之间也存在着某些细微而重要的差别,似乎有进一步划分成亚类的必要。V-ATPase的研究从20世纪70年代后期开始,进展很快。基因克隆和序列分析的结果证明,在进化上,V-和F-ATPAase有比较近的亲缘关系,其结构、生化性质和作用机制类似,它们与P-ATPase有几点主要的差异:(1)在反应过程中不形成共价磷酸化中间物,不受钒酸盐的抑制;(2)都是质子泵ATPase,受阴离子激活;(3)由多个亚基组成的寡聚蛋白。V-ATPase由水溶性的和脂溶性的两部分组成,分子量约400000。水溶性部分由5个亚基组成突向细胞质中,含有催化ATP分解的活性部位。嗜铬颗粒H+-ATPase的可溶性组分中,5个亚基A.B.C.D.E的分子量分别为73000,58000,40000,34000和33000。脂溶性组分的20000和16000的两个亚基中,含有DCCD的结合位点,此外,还有115000和39000两个辅助亚基,前者可与DCCD结合,推测至少这个大的辅助亚基参与质子通道的形成,在笼蛋白衣被小泡的H+-ATPase中也发现辅助亚基,但由于在酶纯化过程中辅助亚基极易脱落,所以目前尚不能确定是否在所有的V-ATPase中都含有辅助亚基。V-ATPase不受寡霉素、叠氮化钠的抑制,不能催化ATP的合成。V-ATPase泵质子进入空泡内,酸化泡浆,为泡内的水解酶提供最适条件,直接或间接影响细胞内的物质分解代谢.在在受体介导的内吞过程中促进受体-配体的离解,对受体的再利用起重要作用。跨膜质子电化学梯度直接影响泡内物质的吸收与贮存。英国Metchell,P.在1961年提出化学渗透假说,解释氧化磷酸化和光合磷酸化的机制,他认为电子传递过程中,由质子泵作用形成的质子电化学梯度(
亚基含有分解ATP的活性部位,β亚基含有ATP的结合位点,此外还有一条称之为ATPase抑制因子的多肽,与F1可逆性的结合。F0组分是与膜结合的脂蛋白,不同来源的F-ATPase复合物中,F0组分的亚基数差别较大,E.Coli的F0有a.b.c三种亚基,c为DCCD结合亚基,每个F0中至少有10个c亚基,和a、b亚基结合形成质子通道。真核生物的Fo通常含有5~8个亚基。全酶具有催化ATP合成与分解双重活性,受寡霉素、叠氮化钠和DCCD的抑制。当ATPase抑制因子与F1结合时,该酶催化ATP合成,反之,催化ATP水解。除机械搅拌外,用EDTA、氯仿处理,也可使F1组分脱落下来,可溶性F1保留催化ATP水解的活性,被称为溶性F1-ATPase,对寡霉素和DCCD不敏感,低温下迅速失活(Coldlabile),其它性质与全酶相似。各种F-ATPase的差异在对抑制剂的敏感性不同。细菌的H+-ATPase对寡霉素不敏感,连接F1和F0的不是OSCP,而是被称之为连接素或结合素(nectin)的蛋白质。叶绿体H+ATPase对寡霉素的敏感性不如线粒体H+对ATPase强,有些甚至完全不敏感。V-ATPase:这类ATPase因存在于真核细胞内的空泡型(Vacuolar)细胞器上而得名,如酵母、真菌的空泡,植物的液泡,动物的溶酶体、笼蛋白衣被小泡(Clathrin Coated Vesicles)、嗜铬颗粒(贮存肾上腺素等激素)、突触小泡(贮存乙酰胆碱)和分泌小泡等。V-ATPase是3类离子转运ATPase中数量最多的一类,它们虽有许多共性,但在它们之间也存在着某些细微而重要的差别,似乎有进一步划分成亚类的必要。V-ATPase的研究从20世纪70年代后期开始,进展很快。基因克隆和序列分析的结果证明,在进化上,V-和F-ATPAase有比较近的亲缘关系,其结构、生化性质和作用机制类似,它们与P-ATPase有几点主要的差异:(1)在反应过程中不形成共价磷酸化中间物,不受钒酸盐的抑制;(2)都是质子泵ATPase,受阴离子激活;(3)由多个亚基组成的寡聚蛋白。V-ATPase由水溶性的和脂溶性的两部分组成,分子量约400000。水溶性部分由5个亚基组成突向细胞质中,含有催化ATP分解的活性部位。嗜铬颗粒H+-ATPase的可溶性组分中,5个亚基A.B.C.D.E的分子量分别为73000,58000,40000,34000和33000。脂溶性组分的20000和16000的两个亚基中,含有DCCD的结合位点,此外,还有115000和39000两个辅助亚基,前者可与DCCD结合,推测至少这个大的辅助亚基参与质子通道的形成,在笼蛋白衣被小泡的H+-ATPase中也发现辅助亚基,但由于在酶纯化过程中辅助亚基极易脱落,所以目前尚不能确定是否在所有的V-ATPase中都含有辅助亚基。V-ATPase不受寡霉素、叠氮化钠的抑制,不能催化ATP的合成。V-ATPase泵质子进入空泡内,酸化泡浆,为泡内的水解酶提供最适条件,直接或间接影响细胞内的物质分解代谢.在在受体介导的内吞过程中促进受体-配体的离解,对受体的再利用起重要作用。跨膜质子电化学梯度直接影响泡内物质的吸收与贮存。英国Metchell,P.在1961年提出化学渗透假说,解释氧化磷酸化和光合磷酸化的机制,他认为电子传递过程中,由质子泵作用形成的质子电化学梯度(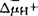 ),
),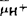 中,所含的渗透能通过ATPase合成ATP。已有许多的事实支持这一假说,并且证明,ΔμH+可直接与耗能过程如溶质跨膜转运偶联。但对泵质子的机理还有待进一步研究。ATPase是生物体内的重要酶之一,研究非常广泛而深入,目前主要对其作机理特别是转运离子的分子机理进行研究。另外,正在研究的古细菌H+-ATPase究竟属于哪一类,是否需要另列新类,还有待进一步研究。【参考文献】:1 Pullman M E.Penefcky H S.Racker E J. Bio! Chem.1960, 232:3322-33292 Mitchell P. Nature .1961, 191:144 - 1483 JosephsonL, Cantley L C. Biochem, 1977.166:4572-20824 Ohkumiy, Anraku Y. Biol Chem, 1981,256: 2079-20825 Wang Yanzhi.Sze H. J Biol Chem,1985,260:10434 - L04436 Hager K M, Mandala S M, Davenport J W, et al. Proc Nail Aead Sci. USA.1986,83:7693 -76977 Nakamoio R K, Slayman C W. J Bioenerg Biomemb. 1989, 21:621-6328 Adachi l.Puopolo K, et al. J Biol Chem, 1990.265:967 - 9739 王延枝,冷楠.武汉大学学报(自然科学版),1991,1∶72~76
中,所含的渗透能通过ATPase合成ATP。已有许多的事实支持这一假说,并且证明,ΔμH+可直接与耗能过程如溶质跨膜转运偶联。但对泵质子的机理还有待进一步研究。ATPase是生物体内的重要酶之一,研究非常广泛而深入,目前主要对其作机理特别是转运离子的分子机理进行研究。另外,正在研究的古细菌H+-ATPase究竟属于哪一类,是否需要另列新类,还有待进一步研究。【参考文献】:1 Pullman M E.Penefcky H S.Racker E J. Bio! Chem.1960, 232:3322-33292 Mitchell P. Nature .1961, 191:144 - 1483 JosephsonL, Cantley L C. Biochem, 1977.166:4572-20824 Ohkumiy, Anraku Y. Biol Chem, 1981,256: 2079-20825 Wang Yanzhi.Sze H. J Biol Chem,1985,260:10434 - L04436 Hager K M, Mandala S M, Davenport J W, et al. Proc Nail Aead Sci. USA.1986,83:7693 -76977 Nakamoio R K, Slayman C W. J Bioenerg Biomemb. 1989, 21:621-6328 Adachi l.Puopolo K, et al. J Biol Chem, 1990.265:967 - 9739 王延枝,冷楠.武汉大学学报(自然科学版),1991,1∶72~76(武汉大学王延枝教授撰;马晓军审)