【孤立子与生命活动】
早在1834年,英国科学家Russell观察到的水渠中出现的奇妙现象:“由两匹马拉着在水渠中急速行进的船,突然停下来,在船前激起一个孤立的水波峰,保持巨大速度和原来形状滚滚向前行进了大约1~2km”。但由于当时科学技术的限制,没有人对这一问题从理论上和实验上作进一步探索。直到60年后的1895年,Kortewwey de Vries才从理论上推导出了描述这种孤立水波的运动方程,即KdV方程式。但人们怀疑KdV方程的解是否稳定。20世纪50年代的Fermi、Pasta和Vlam等人研究的FPU问题,他们将64个质点用非线性弹簧连成一个非线性的振动弦,计算其中能量分布,发现与线性能量均分定理的结果显著不同,由此受到启示:在这个系统中可以出现非线性波。这表明除水系统外,在其它系统中也能观察到孤立波的存在。1965年,美国的Zabrky和Kruskkal用数值方法详细考察了等离子体中孤立波相互碰撞的非线性相互作用过程。1968年,美国的AKNS等人又给出了反散方法求解KdV方程的孤立波的方法。1973年,人们又在冰箱中再现了Rusell的KdV孤立波,从而在世界范围内掀起了“孤波热”。自此以后,人们在大到宇宙,小到基本粒子,复杂到生物体等所有物理系统中都求得了孤立子的存在。
从数学角度讲,它是非线性微分方程的局域性行波解(所谓“局域性”是指解在空间无穷远的处趋于零或确定的常数的情况)。同时,这种局域性行波(或孤立波)在通过相互碰撞后将不消失或碰烂,其波形和速度都不会改变或只有微弱的改变的稳定的孤立波便称便称为孤立子。从物理学角度讲,它是由非线性场所激发的、能量不弥散的、稳定的准粒子。从理论上讲,这种准粒子具有一切物质的属性,如能量、动量、质量、电荷、自旋等特性,并且在运动中或相互碰撞中也不受到破坏;同时,它还遵守无穷多的守恒规律,例如能量、动量、质量守恒定律,所以它具有一切物质或粒子的共性,是物质世界中的一员。但是,由于它是在非线性场中的形成的新型准粒子,因此也具有许多自己“个性”,例如它既具有波动性,又具有粒子性,同时又是遵守经典运动规律的粒子。所谓波动性,因它是真正的一个行波,它可以速度V在时空中传播,但也可以在空间不进行传播,处于静止状态,后者称为非传播孤立子。典型例子是1984年吴汝君和崔洪农与我们观察研究的水槽中的非传播孤立波,它们满足的KdV方程、非线性薛定谔波动方程和非线性Klein-Gordon方程(包括Sine-Gordon方程)等波动方程。说它具有粒子性,是因为它具有如上所述的一切粒子的基本特性,在两个孤立相互碰撞后的规律和一切粒子的碰撞特征与规律是一样的。它比量子力学中讲的微观粒子具有的波粒二象性的物理图像更加具体、更加直接、也更容易理解。它与微观粒子不同的地方,是它遵守经典运动规律,服从牛顿方程式或经典哈密顿方程。这不但可以从它的运动方程中得出,也可从崔洪农与庞小峰等对非传播孤立子所进行的实验中的观察到。这种孤立子受不同外界作用时,可作匀速或匀加速等多种经典运动(这种特性已制成录像带和图片发行)。孤立子的另一个特性是:它是由通常的物质和粒子在色散力和非线性作用力同时存在的环境或场中演化或自陷而成的。例如,现已为日本电信公司作为超高速信号通信的光孤立子是激光光束在非线性介质(如光纤)中的自聚焦而生成的;超导孤立子是超导电子与晶格畸变相分子相互作用生成的;有机分子和蛋白质中的Davydov孤立子或我们在建立的新理论中的孤立子是过剩电子或激子与晶格畸变相互作用“自陷”而成的。由于演变或自陷前的物质的性质不同,以及所受到的性质和所处的环境不一样,或者说物质的性质和所处的环境不一样,使得形成的孤立子的特性千差万别。例如,有些孤立子的质量大到天体中一个星球或云团,有的质量又仅与电子质量相差无几。其能量、电荷等差别就更大了。各种各样的孤立子存在,标志着非线性世界比线性世界更加奇妙,更加复杂,内容更加丰富多彩。另外,从本质上讲,自然界的一切现象基本上是一个非线性问题。所以,非线性问题才是揭示自然界规律的科学,而孤立子问题的研究的意义自然本质重要性也显而易见了。传输生物能量与生物信息的孤立子理论 长期以来,人们对于生命体中生物能量与生物信息传输的微观机制一直未弄清楚,因而对许多生物生理过程(例如肌肉收缩等)的机制及理解模糊不清。然而,人们要去揭示生物的奥秘,发展生物工程,又必须解决这个问题。因为从生物物理的角度来讲,生命过程就是生物能量、物质和信息的变化、协调和统一的过程。其生物组织有秩序地综合运动和相互协调变化则是一切生命的基础。另外,由现代科学得知,生物体是一个具有耗散结构特征和自组织结构的、满足热力学等四定律的、开放的非平衡态热力学系统,它要不断的同外界进行物质、能量和信息交换,以维持生物自组织和耗散结构,即维持生命活动。因此,生物能量与信息传输则是生命活动的最基本、最主要的过程。于是,生物能量和生物信息到底如何传输、以何种形式传输,就成为了生命科学的中心问题。由于生命体不同于一般物体,它是由电子、原子、小分子、生物大分子、细胞或生物组织等多种层次组成的最复杂体系。这种复杂性不但表现为层次多,还在于它是具有自我装配、自我复制、自我调节和自我更新的活的生命体。然而,这些问题的关键都在于生物大分子—-蛋白质和核酸(DNA和RNA)的存在。因此,研究生物能量和信息在蛋白质和核酸中的传输便是此问题的中心了。蛋白质分子系统中生物能量和信息的传输问题引起了A.S.Danydov的极大重视。他和他同事们从60年代末期以来,就一直从事准一维分子系统中孤立激发的研究。Davydov在1979年,将他在一维分子系统中的孤立理论移植到α-螺旋蛋白质分子中,研究了蛋白质分子中的结合能的传输问题。他认为在生物体内,由ATP水解作用等生物过程提供的能量(大约为0.4eV)可以通过蛋白质分子中的过剩电子(excess electorn)或amideI(CO键)振子的振动量子(大约具有0.21eV或1665cm-1的能量)同分子链的结构畸变(或低频声子)相互作用而“自陷”成的孤立子来进行传递。此时,这个孤立子和链的畸变一起携带能量和信息,沿分子链方向传输一段宏观距离而可以保持能量、动量、波形以及其它准粒子特性不变。他将在α-螺旋蛋白质中因集体激发产生的激子和分子链的位移变化的运动方程写成。ihαna=[E0+X(βn+1,a-βn-1,a]αn,a-J(αn+1.a+αn-1,a)+L(αn,a+1-αn.δ-1) (1)Mβna=W(βn+1,a-2βna+βn-1,a)+X(|αn+1.a|2-|αn-1,a|2 (2)而把描述由ATP水解作用等提供的能量而引起的集体激发的尝试波函数写成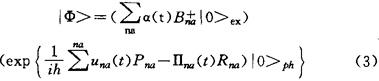
其中E0/h是一个孤立的氨基酸分子中的amide-Ⅰ振子的振动频率,M是一个氨酸分子的质量,J是沿分子链上的两相邻分子间的迁移偶极能,W是氢键的弹性系数,L是两条相邻螺线长链之间的迁移偶极能,X是激光一声子耦合(EPC)常数,它表示了由于氢键链改变一个单位时长度所引起的amide-Ⅰ振子的振动频率的改变。 是激子的产生算符Rna和Pna=MRna分别表示氨基酸分子的纵向位移算符和相应的共轭动量算符。而βna=<Φ|Rna|Φ>.n表示螺旋的圈数,a=1,2,3表示蛋白质分子的3条链,(n,a)规定了一个氨基酸分子。如果对(1)、(2)式作连续性近似,便可得到一个非线性Shuödinger方程(NLSE),马上便可以得到由激子通过与分子链的位移相互耦合而自陷成的Davydov孤立子,这就是传递生物能量和信息的Davydov孤立子机制及理论。在这种机制中,EPC常数X是一个关键因素。许多人的研究表明,产生Davydov孤立子,X应有一个阈值存在。在这个阈值之上,有Davydov孤立子存在,在此值下,则没有。1980年,Kuprievich等人用从头算起法求得X=35PN;后来算得x=(30-60)PN;Pierce算得X=(7-26)PN。Careri等人求得X=62PN;Scott等人用数值计算方法求得X=50PN。X的如此差异是蛋白质分子氢键长度(大约为2.8±0.12Å)取值不一样及所用的方程的近似程度的差别等造成的。另外在氢键长度变化范围内,CO键居于1价与2价之间,因此CO键的强度灵敏地依赖于邻近氢键长度的变化,致使x值也相应改变。现在人们作计算时,对单链蛋白质模型常取X=(20-62)PN,对三链蛋白质模型常取X=(35-62)PN。
是激子的产生算符Rna和Pna=MRna分别表示氨基酸分子的纵向位移算符和相应的共轭动量算符。而βna=<Φ|Rna|Φ>.n表示螺旋的圈数,a=1,2,3表示蛋白质分子的3条链,(n,a)规定了一个氨基酸分子。如果对(1)、(2)式作连续性近似,便可得到一个非线性Shuödinger方程(NLSE),马上便可以得到由激子通过与分子链的位移相互耦合而自陷成的Davydov孤立子,这就是传递生物能量和信息的Davydov孤立子机制及理论。在这种机制中,EPC常数X是一个关键因素。许多人的研究表明,产生Davydov孤立子,X应有一个阈值存在。在这个阈值之上,有Davydov孤立子存在,在此值下,则没有。1980年,Kuprievich等人用从头算起法求得X=35PN;后来算得x=(30-60)PN;Pierce算得X=(7-26)PN。Careri等人求得X=62PN;Scott等人用数值计算方法求得X=50PN。X的如此差异是蛋白质分子氢键长度(大约为2.8±0.12Å)取值不一样及所用的方程的近似程度的差别等造成的。另外在氢键长度变化范围内,CO键居于1价与2价之间,因此CO键的强度灵敏地依赖于邻近氢键长度的变化,致使x值也相应改变。现在人们作计算时,对单链蛋白质模型常取X=(20-62)PN,对三链蛋白质模型常取X=(35-62)PN。
 的公式中得到证实。在T=T0时,孤立子变成一个无限宽的扩展态(这不是孤立子状态)。对于三螺线蛋白质模型来讲,T0=100~120K,即Davydov孤立子也不可能在300K的温度下存在下去。基于这些问题,在1989年丹麦召开的专门讨论Davydov孤立子的国际会议上,代表们一致认为Davydov理论是一个十分粗糙和不完善理论,需要改进和发展。Takeno等人建议关于白陷量子振动能量的Davyda描述应修改为表示允许振动量子数随时间而改变。它可以通过把迁移偶极算符
的公式中得到证实。在T=T0时,孤立子变成一个无限宽的扩展态(这不是孤立子状态)。对于三螺线蛋白质模型来讲,T0=100~120K,即Davydov孤立子也不可能在300K的温度下存在下去。基于这些问题,在1989年丹麦召开的专门讨论Davydov孤立子的国际会议上,代表们一致认为Davydov理论是一个十分粗糙和不完善理论,需要改进和发展。Takeno等人建议关于白陷量子振动能量的Davyda描述应修改为表示允许振动量子数随时间而改变。它可以通过把迁移偶极算符 改变成为
改变成为 来得到。Lindenberg和MigiLiori等人拥护这种观点,但得到的结果并不理想。Wang等人改进计算方法出发,也未得到好的结果。看来只动“小手术”改进计算方法等无法收到好的效果。必须对蛋白质分子中的集体激发特性进行重新认识和重新研究,才可能收到好的效果。庞小峰等人认真分析了Davydov理论之所以不适合于生物蛋白质分子系统的重要原因是,Davydov理论是把他多年研究的一般分子系统的孤立子理论机械进移植到蛋白质分子中所造成的。但生物大分子的蛋白质是不同于一般分子系统的,它是一类生物自组织系统。因此在建立描述该体系中集体激发的哈密顿函数和波函数时,必须考虑这个基本特性,不能机械照搬。我们在分析由ATP水解作用等提供的能量而引起的蛋白质分子结构畸变和局域性涨落的特点后,给出了一个较为全面反映蛋白质分子的动力学特征的哈密顿量
来得到。Lindenberg和MigiLiori等人拥护这种观点,但得到的结果并不理想。Wang等人改进计算方法出发,也未得到好的结果。看来只动“小手术”改进计算方法等无法收到好的效果。必须对蛋白质分子中的集体激发特性进行重新认识和重新研究,才可能收到好的效果。庞小峰等人认真分析了Davydov理论之所以不适合于生物蛋白质分子系统的重要原因是,Davydov理论是把他多年研究的一般分子系统的孤立子理论机械进移植到蛋白质分子中所造成的。但生物大分子的蛋白质是不同于一般分子系统的,它是一类生物自组织系统。因此在建立描述该体系中集体激发的哈密顿函数和波函数时,必须考虑这个基本特性,不能机械照搬。我们在分析由ATP水解作用等提供的能量而引起的蛋白质分子结构畸变和局域性涨落的特点后,给出了一个较为全面反映蛋白质分子的动力学特征的哈密顿量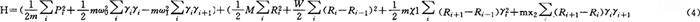
这里不但考虑了低频声学声子的作用,而且也考虑了纵向光学声子的作用,同时还考虑了amide-I振子与所在氨基酸分子及相邻氨基酸分子的振动量子(声子)之间的相互作用。并且我们还论证了这个具有自组织特征的蛋白质中所产生的集体激发所具的相干特性,因而我们应用了一个新的准相干态波函数(即我们的波函数),而不是Davydov波函数来描述这种状态,即将该体系的波函数表示为
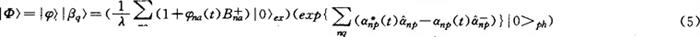
再采用新方法,得出了与Davydov理论迥然不同的结果。例如其孤立子的能量为(1.49~4.46)×10-3eV,与实验值1.86×10-3eV相当接近,但Davydov理论得到的孤立子能量为1.4×10-4eV,与实验值相差甚远。我们还得到激子速度vg=103m·s-1。声速vaq为5×103m·s-1≤Vaq≤104m·s-1,与实验值(3.6-44.8)×103m·s-1接近。但Davydov理论得到的vaa=100m·s-1,与实验值相差较远。计算了由新孤立子运动产生的比热也与实测值吻合。我们又研究了这类新孤立子的温度效应,发现了这类新振幅虽然随温度而改变,但在310K时,这类孤立子仍是稳定的。单就温度影响来讲,在310K时仅比10K时增加2~3倍。但若又考虑由温度不等于零而引起的分子链的非线性振动效应对孤立子振幅引起的减少的抵消作用后,这孤立子在310K的振幅变化不大,因而能保持稳定。临界温度可达348~359K。庞小峰等还计算这类孤立子在310K时的寿命可达10-9~10-8s,与Princeton大学R.Austin等人实测结果10-10s很接近,在这段时间内,它可通过大约104个氨基酸分子。这表明这类孤立子够也承担起自己的生物功能,在生物学过程中是非常有用的。另外又论证了这类孤立子的量子特性,非线性振动对它的影响,超声速运动特征,自发辐射效应,拉曼散射以及穆斯堡尔效应等问题,都获得了较合理的结果,说明了他们的理论是正确的,适合蛋白质分子系统。
关于蛋白质分子的生物能量由孤立子传递的实验研究已由Webb和Careri等人(1984)对与它的结构完全相似的[CH3CONHC6H5]x分子晶体中的拉曼光谱和红外谱测试和Faum等人对蛋白质分子所作的自由电子激光脉冲测试以及BobKnkK等人的微微秒泵-探针实验测定工作中所证实。看来,以上的理论是正确的。核酸中的孤立子及其生物功能 对于另一类生物大分子——核酸(DNA和RNA)则是生物遗传信息的负载者,包括人在内的生物之所以能一代一代相传下去,就是由DNA分子的解链和复制功能引起的。并且它还是制造蛋白质的模板。然而人们一直未弄清楚这个复制过程是如何完成的。因而对生物遗传与变异及癌症等疾病的原因一直未弄清楚。实验测定复制叉的移动速度,对原核细胞约为每秒1000核苷酸。对于真核细胞约为每秒100核苷酸其的速度大约为10-5cm·s-1的数量级。如果我们把解链过程看成机械的螺旋转动,则DNA转动速度大约应为10~100r/s。这么大的速度,对DNA的运动来讲是无法理解的。但若把DNA的解链看成一种量子力学的构象改变,运用微观动力学理论可以估计出复制叉的移动速度约为105cm·s-1,大大高于实验值。若放弃微观动力学观点,采用统计力学的途径,可得到合理的复制叉速度,但又难于解释链与复制的高度有序性。看来DNA的解链与复制是一个非常复杂的问题。但我们最近利用DNA的孤立子运动和分形理论可能对这个问题有较合理的解释。关于DNA中的孤立子运动,最早由S.N.Englander等人(1980)研究过,以后S.Yomosa(1989)等人及庞小峰对B-DNA的平面基转子模型的孤立子理论也作过广泛、深入的研究。近似认为DNA的碱基(C.T.A.G)是一个平面转子。双螺旋中的两个转子在ATP水解作用和外界刺激提供的能量作用下,可分别绕着螺旋轴转动(其转角为φnφn),从而改变了DNA的构象,引起互补碱基之间的氢键能,相邻碱基之间的堆积能和螺旋状扭转能,碱基集团之间的范德瓦斯相互作用能和Frōhlish吸收能等的变化。考虑这些效应,我们把第n个互补碱基的运动方程表示为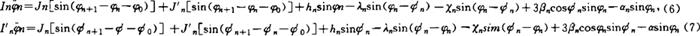
(西南民族学院庞小峰撰)