【突变体在植物激素研究中的应用】
拼译:use of mulants on phytohormone research
自从1956年Phinny最早发现了缺少GA的玉米矮生突变体以来,到目前为止,人们已经获得了许多突变体,它们对研究植物激素的生物合成途径,生长发育的激素控制以及在分子水平上研究激素的机理等,有着特殊的意义,有些突变体还有商业价值。
植物激素突变体可以分为两类:(1)影响激素水平的合成突变体,(2)影响激素反应的敏感性突变体。最常见的合成突变体是阻断激素合成的某些步骤的突变体,如缺少GA1的矮生豌豆和玉米,缺少ABA的番茄萎蔫突变体,藓(Physcomitrellapatens)是细胞分裂素过量产生(over producing)的突变体等。绝大多数突变体是属于敏感性的类型,它们影响激素的亲和力或受体分子,它们在表型上与合成突变体没有什么差别。GA突变体1.玉米。Phinney最早证明玉米节间伸长是受激素基因控制的。玉米矮生的表型可以从幼苗到成熟的不同时期表现出来。影响玉米高度的突变体有30多个,涉及5个非等位基因d1,d2,d3,d5,an1。这些突变体的表型特征是在外加GA3后,植物恢复正常高度。矮化的植株比正常植株体内GA活性小。因此认为这些突变体的矮化,是因为GA生物合成途径中不同的步骤受到抑制的结果。此外,还有另外5个非等位基因的突变体D8,pe1,mi2,na1和na2,它们对外加GA3反应很小或无反应。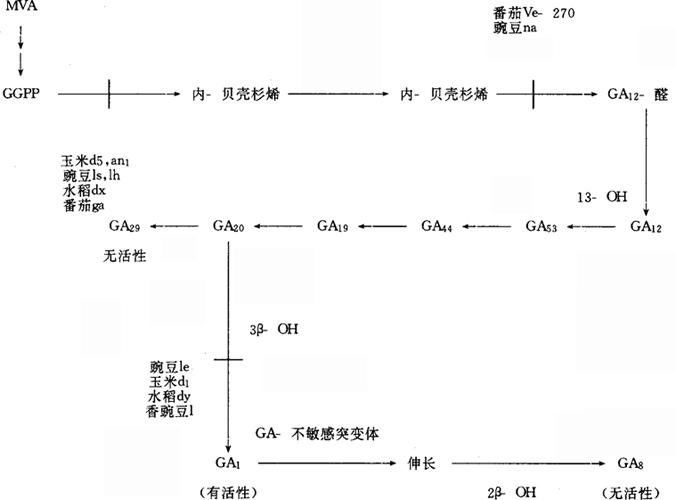
图1 GA生物合成的13-羟基化途径及合成突变体可能的作用位点
GA合成的途径是从甲瓦龙酸(MVA)起始。二甲烯丙基焦磷酸酯(DAMPP)与三分子异戊烯基焦磷酸(IPP)缩合,形成 牛儿
牛儿 牛儿基焦磷酸(GGPP),环化形成珂巴基焦磷酸(CPP),进一步环化形成内贝壳杉烯,然后形成GA12醛。GA12醛是所有GA的前体。玉米中从GA12醛开始,通过13-羟基化,产生GA53、GA44、GA19、GA20、GA1。此外,这个途径的副代谢产物还有GA17、GA29、GA8,它们都是无活性的。现已证明玉米的伸长与GA1的控制有密切关系,而与植株中总体GA无关。d5阻碍CPP环化成为内贝壳杉烯。这个步骤是由内贝壳杉烯合成酶所催化的。正常玉米幼苗茎的无细胞抽提物可以合成内贝壳杉烯,但d5幼苗的抽提物,无论以甲瓦龙酸为底物还是以GGPP或CPP为底物,合成的贝壳杉烯速度只有正常幼苗中1/5。d1的基因阻碍GA20→GA1,这个步骤是3β-羟基化。这个基因的作用位点可以从下列实验得到证明。(1)用[13C,3H]GA20双标记物供给正常幼苗,产生GA29和GA1标记物,而d1幼苗只产生GA29及其分解物,不产生GA1。(2)在d1幼苗内GA1(包括其无活性代谢物GA8)比正常幼苗中少2%。(3)d1幼苗中GA20(无活性代谢物GA29)的积累比正常幼苗中多10倍。d2可能控制7C-氧化步骤即GA12醛→GA53。因为在幼苗上加GA53醛没有活性,加GA53则叶鞘伸长。d3可能控制13-C-羟基化步骤,因为幼苗加GA12-醛不表现活性,而加GA53醛则表现活性。现在发现了约20个以上GA不敏感型的玉米突变体,如D8。它的表型特点是叶片短而宽,卷曲,节间短,雌雄穗小,花粉发育好。这组突变体对外加GA无反应,说明它们的作用是在GA1合成以后的步骤。虽然,从GA被受体接受到伸长所发生的事件还不清楚,但用这类突变体可以解释这些步骤。例如,GA可以改变微管和微丝的取向,这对细胞的伸长是十分重要的。突变体影响伸长可能是改变了表皮细胞的长度或每个节间的细胞数目。所以利用敏感突变体研究伸长的生理过程是有意义的。2.豌豆。Brian(1957)最早研究豌豆节间伸长基因。但在60~70年代并不了解高生与矮生豌豆在GA类似物上有什么区别。近来证明豌豆的矮化也受GA1基因控制。高生型(Le)豌豆体内有GA1,在光下生长的矮生型(le)则没有GA1。le阻碍GA20→GA1的3β-羟基化步骤。在Le中,[3H,13C]GA20可以转变为GA1、 GA8或GA29,le中[3H,13C]GA20不能转变为GA1,只产生少量GA8。这表明le是一个渗漏突变体(leakymutant)。这些资料证明GA1是le豌豆中唯一影响伸长的激素,就如d1在玉米中,dy在水稻中一样。在豌豆中除le外,还研究了7个非等位基因,它们是na,lh,ls,la,cry,lk,lm,有些是GA敏感突变体。豌豆突变体lv的反应与光有关。在暗中或远红光中lv与正常野生种Lv在节间长度上没有区别,在连续红光或白光下,lv的节间缩短,但它们的GA1或GA20水平在光下并没有改变。因此认为可能是光通过光敏素系统影响GA的敏感性而不是影响GA的水平而控制伸长。除对玉米、豌豆的突变体做了较详细的研究外,近年来也用突变体研究了牵牛、水稻、小麦、番茄、拟南芥和四季豆的有关伸长基因。ABA突变体 已经研究的ABA合成突变体有番茄萎蔫突变体flc,sil,nol,土豆萎蔫突变体dp,豌豆萎蔫突变体wil,拟南芥萎蔫突变体aba等,这些突变体导致ABA的水平降低。玉米vp-5,w-3,vp-7突为体是由于阻碍类胡萝卜素合成而降低ABA水平。现在认为ABA合成途径是:(1)直接的C15途径,即法尼基焦磷酸(FPP)环化及氧化;(2)间接的C40途径,从紫黄质分解。目前,更多的研究者支持后者。例如,玉米突变体阻碍类胡萝卜素生物合成,并导致ABA水平降低,这暗示ABA合成需要类大胡萝卜素的中间产物。利用类胡萝卜素合成的抑制剂支持这个结论。如果C40中间产物断开得到ABA的C15前体,则将会形成两种副产物,C25(一次断裂)或C10(两次断裂)。Taylor用缺少ABA的番茄突变体,发现了一个C10产物,2,7-二甲基-2,4-辛二烯酸(ODA)。在flc和sit突变体中ODA水平比野生型中高,这表明在C40前体物断裂后,flc和sil中ABA合成受阻。在nol突变体中OAD水平比野生型中低,这表明not中ABA合成的抑制是在C40前体物断裂后,flc和sit中ABA合成受阻。在not突变体中OAB水平比野生型中低。这表明not中ABA合成的抑制是在C40断裂过程时或断裂以前,植株受到水胁迫时更是如此。Lindforth等证明在flc和sit中以如同ODA一样的方式积累2-反式-ABA-醇,并达到足以降低ABA水平的近似量。这证明2-反式ABA-醇为ABA合成的中间产物。Parry等研究了紫黄质分解的产物,14C-叶黄氧化素,转化成ABA。他们发现在野生和not突变体中发生转化,而在flc,sit中只限于位置上的转化。这些结果与Lindforth的结果是一致的。但是详细的中间产物与这3种基因的作用仍不清楚。ABA萎蔫突变体的表型特点是气孔不关闭,植株萎蔫。突变体的叶片比野生型失水快。它们的叶片离体后严重失水,对干旱的反应是不积累ABA。同时,虽然在水分胁迫下,气孔导度仍很高。土豆突变体在缺水5d时,基部叶片导度仍为0.9~1.0cm·s-1,此时叶片已萎蔫。而同样萎蔫程度的野生型叶片(13d不给水),导度下降到0.1~0.4cm·s-1。从拟南芥、大麦、水蕨类Ceratopteris等植物中分离得到了ABA不敏感的突变体。这些突变体对外加ABA不表现敏感性。有些突变体只影响一部分敏感性。如拟南芥abi-3只降低种子休眠,而“cool”大麦突变体用ABA处理后不关闭气孔。这些结果证明,对ABA的各种作用:种子休眠、幼苗生长、气孔关闭来说,至少有某些共同的步骤。在拟南芥中至少有2个作用位点。突变体似乎只影响某一个生理过程,所以它是受体突变体。生长素突变体 虽然在生长素合成突变体方面做了很多研究,但到目前为止,还没有检测到高等植物的生长素合成突变体。原因可能是这种基因是致死的,或是对它们表现形式还缺乏了解。Blonstein等(1988)从Nicotiana plumbaginifolia原生质体中发现一个突变细胞系,是生长素缺陷型突变体。它们的幼茎即使嫁接到野生的烟草上,也不能获得再生植株。它的生长呈丛生状,顶端优势弱,叶子生长慢,叶片干枯前,老叶上出现褐色斑点,并发生脱落死亡,证明它是生长素缺陷型。藓Physcomitrella patens从原叶状体转变成茎丝体,需要生长素和细胞激动素。有的突变体不能完成这个转变,可能是生长素强烈分解的结果。巨型浮萍(Lemna gibba)变种是由于游离IAA的增加,但其基因控制和IAA改变的意义不太明确。拟南芥有色氨酸合成突变体trpl-1,它生长缓慢,叶卷曲,丝生,花不育,其体内IAA含量低。用遗传操作手段可以通过转基因植物提高内源生长素水平。例如,致癌农杆菌T-DNA上合成生长素的基因已经被分离出来,并转到矮牵牛中,在启动子控制下得到表达。结果这些植物中IAA含量比野生型产长10倍,植物顶端优势明显,节间变长,叶子卷曲,木质部和韧皮部增加,能单性结实,这些性状是因为IAA过量产生所致。用这种转基因植物研究生长素过量产生在特定组织中的作用有广阔前景。生长素敏感突变体比合成突变体更为普遍。番茄横向重力性(dgt)突变体就是一例。对外加IAA不敏感,下胚轴不伸长,不产生乙烯。认为是它的生长素原初作用位点受到影响所致。但现有资料还不能说明这一点。这个突变体的表现也与乙烯有关,所以它是一个乙烯突变体。近来King等研究了Nicotiana plumbaginifolia化学诱变突变体。培养基中加入20μmol·L-1NAA,野生型生长受抑制,根、芽畸形,而抗生长素突变体生长正常。在加20μmol·L-1BAP培养基上,抗细胞激动素突变体茎叶正常。此外,还从这种植物突变体的原生质体中筛选出生长素自养型(auxin auxotroph)和温度敏感型的个体。在加NAA的培养基上,野生型生长正常.而生长素自养型突变体则生长受抑制。这说明它们生长不依赖于生长素。生长素自养型突变体和野生型分别培养在33℃+生长素,33℃-生长素,26℃+生长素,26℃-生长素的条件下。在33℃+生长素处理中,突变体叶子变褐或退色,芽生长较差。说明这个突变体对温度敏感,而且其生长不需生长素。用筛选出的N.plumbaginifolia突变体研究IAA代谢,结果表明在野生型和突变体细胞中,IAA半衰期为20min。IAA最初的代谢产物为IAA-天冬氨酸和IAA-葡萄糖酯。当培养基中不含IAA时,野生型和突变体二者在代谢上表现不同。突变体中IAA-天冬氨酸代谢快,半衰期27min,而野生型中为9.5h。细胞激动素突变体 Physcomitrella patens是细胞激动素过量产生的突变体,它的iPA和ZT含量比野生型高100倍。配子托数量多,就像野生型用细胞激动素处理一样。表明细胞激动素对配子托的形成有重要作用。体细胞杂交证明至少有3个基因控制着细胞激动素的产生。也有细胞激动素缺失突变体,它不产生配子托,加细胞激动素就可以产生。这可以用来研究藓的细胞激动素合成。可惜在高等植物中还没有发现细胞激动素合成突变体。但从致癌农杆菌中已分离出细胞激动素合成基因,并在高等植物中得到表达。
牛儿基焦磷酸(GGPP),环化形成珂巴基焦磷酸(CPP),进一步环化形成内贝壳杉烯,然后形成GA12醛。GA12醛是所有GA的前体。玉米中从GA12醛开始,通过13-羟基化,产生GA53、GA44、GA19、GA20、GA1。此外,这个途径的副代谢产物还有GA17、GA29、GA8,它们都是无活性的。现已证明玉米的伸长与GA1的控制有密切关系,而与植株中总体GA无关。d5阻碍CPP环化成为内贝壳杉烯。这个步骤是由内贝壳杉烯合成酶所催化的。正常玉米幼苗茎的无细胞抽提物可以合成内贝壳杉烯,但d5幼苗的抽提物,无论以甲瓦龙酸为底物还是以GGPP或CPP为底物,合成的贝壳杉烯速度只有正常幼苗中1/5。d1的基因阻碍GA20→GA1,这个步骤是3β-羟基化。这个基因的作用位点可以从下列实验得到证明。(1)用[13C,3H]GA20双标记物供给正常幼苗,产生GA29和GA1标记物,而d1幼苗只产生GA29及其分解物,不产生GA1。(2)在d1幼苗内GA1(包括其无活性代谢物GA8)比正常幼苗中少2%。(3)d1幼苗中GA20(无活性代谢物GA29)的积累比正常幼苗中多10倍。d2可能控制7C-氧化步骤即GA12醛→GA53。因为在幼苗上加GA53醛没有活性,加GA53则叶鞘伸长。d3可能控制13-C-羟基化步骤,因为幼苗加GA12-醛不表现活性,而加GA53醛则表现活性。现在发现了约20个以上GA不敏感型的玉米突变体,如D8。它的表型特点是叶片短而宽,卷曲,节间短,雌雄穗小,花粉发育好。这组突变体对外加GA无反应,说明它们的作用是在GA1合成以后的步骤。虽然,从GA被受体接受到伸长所发生的事件还不清楚,但用这类突变体可以解释这些步骤。例如,GA可以改变微管和微丝的取向,这对细胞的伸长是十分重要的。突变体影响伸长可能是改变了表皮细胞的长度或每个节间的细胞数目。所以利用敏感突变体研究伸长的生理过程是有意义的。2.豌豆。Brian(1957)最早研究豌豆节间伸长基因。但在60~70年代并不了解高生与矮生豌豆在GA类似物上有什么区别。近来证明豌豆的矮化也受GA1基因控制。高生型(Le)豌豆体内有GA1,在光下生长的矮生型(le)则没有GA1。le阻碍GA20→GA1的3β-羟基化步骤。在Le中,[3H,13C]GA20可以转变为GA1、 GA8或GA29,le中[3H,13C]GA20不能转变为GA1,只产生少量GA8。这表明le是一个渗漏突变体(leakymutant)。这些资料证明GA1是le豌豆中唯一影响伸长的激素,就如d1在玉米中,dy在水稻中一样。在豌豆中除le外,还研究了7个非等位基因,它们是na,lh,ls,la,cry,lk,lm,有些是GA敏感突变体。豌豆突变体lv的反应与光有关。在暗中或远红光中lv与正常野生种Lv在节间长度上没有区别,在连续红光或白光下,lv的节间缩短,但它们的GA1或GA20水平在光下并没有改变。因此认为可能是光通过光敏素系统影响GA的敏感性而不是影响GA的水平而控制伸长。除对玉米、豌豆的突变体做了较详细的研究外,近年来也用突变体研究了牵牛、水稻、小麦、番茄、拟南芥和四季豆的有关伸长基因。ABA突变体 已经研究的ABA合成突变体有番茄萎蔫突变体flc,sil,nol,土豆萎蔫突变体dp,豌豆萎蔫突变体wil,拟南芥萎蔫突变体aba等,这些突变体导致ABA的水平降低。玉米vp-5,w-3,vp-7突为体是由于阻碍类胡萝卜素合成而降低ABA水平。现在认为ABA合成途径是:(1)直接的C15途径,即法尼基焦磷酸(FPP)环化及氧化;(2)间接的C40途径,从紫黄质分解。目前,更多的研究者支持后者。例如,玉米突变体阻碍类胡萝卜素生物合成,并导致ABA水平降低,这暗示ABA合成需要类大胡萝卜素的中间产物。利用类胡萝卜素合成的抑制剂支持这个结论。如果C40中间产物断开得到ABA的C15前体,则将会形成两种副产物,C25(一次断裂)或C10(两次断裂)。Taylor用缺少ABA的番茄突变体,发现了一个C10产物,2,7-二甲基-2,4-辛二烯酸(ODA)。在flc和sit突变体中ODA水平比野生型中高,这表明在C40前体物断裂后,flc和sil中ABA合成受阻。在nol突变体中OAD水平比野生型中低,这表明not中ABA合成的抑制是在C40前体物断裂后,flc和sit中ABA合成受阻。在not突变体中OAB水平比野生型中低。这表明not中ABA合成的抑制是在C40断裂过程时或断裂以前,植株受到水胁迫时更是如此。Lindforth等证明在flc和sit中以如同ODA一样的方式积累2-反式-ABA-醇,并达到足以降低ABA水平的近似量。这证明2-反式ABA-醇为ABA合成的中间产物。Parry等研究了紫黄质分解的产物,14C-叶黄氧化素,转化成ABA。他们发现在野生和not突变体中发生转化,而在flc,sit中只限于位置上的转化。这些结果与Lindforth的结果是一致的。但是详细的中间产物与这3种基因的作用仍不清楚。ABA萎蔫突变体的表型特点是气孔不关闭,植株萎蔫。突变体的叶片比野生型失水快。它们的叶片离体后严重失水,对干旱的反应是不积累ABA。同时,虽然在水分胁迫下,气孔导度仍很高。土豆突变体在缺水5d时,基部叶片导度仍为0.9~1.0cm·s-1,此时叶片已萎蔫。而同样萎蔫程度的野生型叶片(13d不给水),导度下降到0.1~0.4cm·s-1。从拟南芥、大麦、水蕨类Ceratopteris等植物中分离得到了ABA不敏感的突变体。这些突变体对外加ABA不表现敏感性。有些突变体只影响一部分敏感性。如拟南芥abi-3只降低种子休眠,而“cool”大麦突变体用ABA处理后不关闭气孔。这些结果证明,对ABA的各种作用:种子休眠、幼苗生长、气孔关闭来说,至少有某些共同的步骤。在拟南芥中至少有2个作用位点。突变体似乎只影响某一个生理过程,所以它是受体突变体。生长素突变体 虽然在生长素合成突变体方面做了很多研究,但到目前为止,还没有检测到高等植物的生长素合成突变体。原因可能是这种基因是致死的,或是对它们表现形式还缺乏了解。Blonstein等(1988)从Nicotiana plumbaginifolia原生质体中发现一个突变细胞系,是生长素缺陷型突变体。它们的幼茎即使嫁接到野生的烟草上,也不能获得再生植株。它的生长呈丛生状,顶端优势弱,叶子生长慢,叶片干枯前,老叶上出现褐色斑点,并发生脱落死亡,证明它是生长素缺陷型。藓Physcomitrella patens从原叶状体转变成茎丝体,需要生长素和细胞激动素。有的突变体不能完成这个转变,可能是生长素强烈分解的结果。巨型浮萍(Lemna gibba)变种是由于游离IAA的增加,但其基因控制和IAA改变的意义不太明确。拟南芥有色氨酸合成突变体trpl-1,它生长缓慢,叶卷曲,丝生,花不育,其体内IAA含量低。用遗传操作手段可以通过转基因植物提高内源生长素水平。例如,致癌农杆菌T-DNA上合成生长素的基因已经被分离出来,并转到矮牵牛中,在启动子控制下得到表达。结果这些植物中IAA含量比野生型产长10倍,植物顶端优势明显,节间变长,叶子卷曲,木质部和韧皮部增加,能单性结实,这些性状是因为IAA过量产生所致。用这种转基因植物研究生长素过量产生在特定组织中的作用有广阔前景。生长素敏感突变体比合成突变体更为普遍。番茄横向重力性(dgt)突变体就是一例。对外加IAA不敏感,下胚轴不伸长,不产生乙烯。认为是它的生长素原初作用位点受到影响所致。但现有资料还不能说明这一点。这个突变体的表现也与乙烯有关,所以它是一个乙烯突变体。近来King等研究了Nicotiana plumbaginifolia化学诱变突变体。培养基中加入20μmol·L-1NAA,野生型生长受抑制,根、芽畸形,而抗生长素突变体生长正常。在加20μmol·L-1BAP培养基上,抗细胞激动素突变体茎叶正常。此外,还从这种植物突变体的原生质体中筛选出生长素自养型(auxin auxotroph)和温度敏感型的个体。在加NAA的培养基上,野生型生长正常.而生长素自养型突变体则生长受抑制。这说明它们生长不依赖于生长素。生长素自养型突变体和野生型分别培养在33℃+生长素,33℃-生长素,26℃+生长素,26℃-生长素的条件下。在33℃+生长素处理中,突变体叶子变褐或退色,芽生长较差。说明这个突变体对温度敏感,而且其生长不需生长素。用筛选出的N.plumbaginifolia突变体研究IAA代谢,结果表明在野生型和突变体细胞中,IAA半衰期为20min。IAA最初的代谢产物为IAA-天冬氨酸和IAA-葡萄糖酯。当培养基中不含IAA时,野生型和突变体二者在代谢上表现不同。突变体中IAA-天冬氨酸代谢快,半衰期27min,而野生型中为9.5h。细胞激动素突变体 Physcomitrella patens是细胞激动素过量产生的突变体,它的iPA和ZT含量比野生型高100倍。配子托数量多,就像野生型用细胞激动素处理一样。表明细胞激动素对配子托的形成有重要作用。体细胞杂交证明至少有3个基因控制着细胞激动素的产生。也有细胞激动素缺失突变体,它不产生配子托,加细胞激动素就可以产生。这可以用来研究藓的细胞激动素合成。可惜在高等植物中还没有发现细胞激动素合成突变体。但从致癌农杆菌中已分离出细胞激动素合成基因,并在高等植物中得到表达。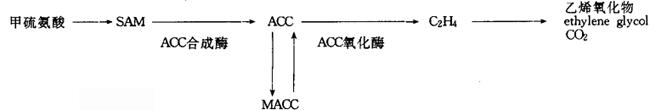
图2 乙烯的生物合成与代谢
乙烯突变体 乙烯合成途径被许多酶所催化,ACC合成酶是限速酶,它对乙烯合成很重要(图2)。Zobel(1973)发现了一个番茄缺乙烯突变体dgt(diageotropica),其表型特点是茎与根水平生长(横向重力),茎细而坚硬,叶呈深绿色,偏上性,初生根和不定根不分枝。外加乙烯(<0.005μl·L-1)后,突变体生长正常。此突变体对IAA反应不敏感,即加IAA后,不能使乙烯的合成增加。由于IAA可以促进ACC合成酶活性,促进乙烯生成,因此认为dgt是一个乙烯合成突变体。此突变体及其等基因亲本VFN8,适用于研究IAA诱导的乙烯生成,及其在控制植物形态学和生理学方面的相互关系。也有人用它研究干旱条件下乙烯生成。dgt组织中的酶活性可以因为受伤或厌氧条件而提高,故有人认为也许基因是与IAA受体有关,而不是与乙烯合成途径本身有关。成熟、衰老、脱落突变体 已筛选出了几个与番茄成熟性状有关的突变体。rin,Nr,nor,alc。它们的特性如表1。这些果实不易变软的特性,特别适合做商品,运输方便,耐贮藏。可随市场需要而催熟出售。表1 番茄成熟突变体的特性
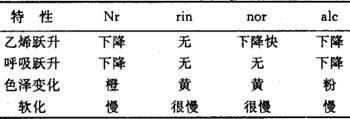
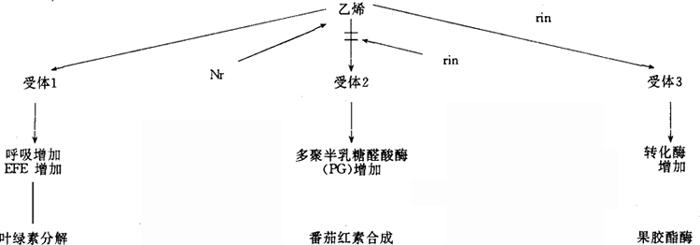
图3 rin和Nr突变作用的多元受体机理
最近报道,用拟南芥抗乙烯突变体er研究乙烯对叶片衰老的作用。er是抗乙烯突变体,它的特性是在空气中含15μl·L-1乙烯中放3d,叶能保持绿色,而野生型则衰老变黄。er的离体叶圆片,无论在光下或暗中,叶绿素分解都慢。光下突变体乙烯合成更快。BA减慢野生型和突变体叶绿素分解,ABA则加速分解。ABA处理能增加突变体乙烯产生,但AOA阻止乙烯合成时,ABA也使突变体衰老变黄,说明ABA促进衰老不是通过乙烯合成的增加,乙烯只是叶衰老的一个因子。植物激素突变体的研究前景广阔。这一领域的工作在我国基本上是个空白点。【参考文献】:1 Reid JB. Phytohormone mutants in plant research. J Plant Growth Reg,1990,9:97~1112 Reid JB. The genetic control of growth via hormones. In: Davies PJ ed. Plant hormones and their role in plant growth and development. Martinus Nijhoff, Wordrecht, The Nederland,1987. 318~3403 Reid JB,Davies PJ. The genetics and physiology of gibbere-llin sensilivity mutants in peas. In:Karssen CM,Van Loon, Vreugdenhil (eds). Progress in plant growth regulation, 1992.214~2254 Taylor I B. ABA deficient mutants. In: Thomas H. Grierson D(eds). Developmental mutants in higher plants. Cambridge University Press,Cambridge, 1987.197~2215 Quarrie SA. Use of genotype in endogenous abscisic acid le-vels in studies of physiology and development. In: Hoad(中国农业大学农业生物学院韩碧文教授撰)