| 释义 |
【漆酶】
拼译:lacccase
漆酶(EC1.10.3.2是含铜氧化还原酶,主要有漆树漆酶和真菌漆酶两类,分别来源于漆树液(生漆)和真菌。研究漆酶的结构与功能、金属铜离子的作用与配位环境、催化反应、氧化还原和电子传递过程等,不仅为人们了解天然体系、探索生命现象提供信息,而且对天然资源的开发、应用都具有重要意义。 1883年,日本吉田首次从日本生漆(即中国生漆)中发现漆酶,但他没有深入研究。后来,伯查德(Bertrand)和凯林(Keilin)分别从越南生漆和中国生漆中分离出漆酶,发现它能催化氧化漆酚、邻苯二酚、对苯二酚、连苯三酚等多元酚。1897年,拉波德(Laborde)发现真菌类植物中有一种酶具有漆树酶一样的催化活性,因而称真菌漆酶。此外,还在紫花苜蓿、毒莴苣、马铃薯、毛地黄、香蕉、咖啡豆、苹果等植物中发现漆酶。不同来源的漆酶化学组成也有差别(表1)。表1 漆酶的分子量、含糖量和含铜量 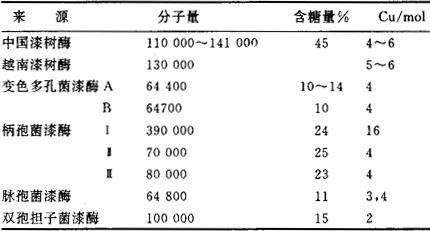
按光谱和磁学性质可以将漆酶中的铜离子(Cu2+)分为3类:Ⅰ型铜离子和Ⅱ型铜离子各1个,Ⅲ型铜离子两个。Ⅰ型铜离子呈蓝色,在λ614nm处有特征吸收;Ⅱ型铜离子非蓝色,没有特征吸收光谱。Ⅲ型铜离子是偶合的离子对(Cu2+-Cu2+),在λ330nm处有宽的吸收峰。Ⅰ型铜离子和Ⅱ型铜离子呈顺磁性,可用ESR检测,Ⅲ型铜反磁性,一般条件下ESR检测不出。Ⅱ型铜离子可选择性地除去,得部分脱辅基漆酶,即T2D(Type Ⅱ Cudepleted)漆酶。T2D漆酶无催化活性,用Cu+离子重建可使其恢复天然酶活性。Ⅰ型铜离子可用Hg2+或Co2+离子取代,得部分取代的漆酶,它的催化活性比天然漆酶低。由于漆酶的含糖量较高,难以得到X-射线分析用的漆酶单晶,它的三维空间结构还不清楚。根据光谱、磁学性质和实验事实推测,漆酶中铜的配位结构为Ⅰ型铜离子分别与Cys、Met中的两个硫、两个His中的氮原子配位,形成变形的四面体。Ⅱ型铜离子是四方配位,分别与2个或3个His中的氮和两个或一个氧原子配位,氧化型的Ⅱ型铜离子含氧配体可能是水或羟基。Ⅲ型铜离子中的2个铜分别与3个His中的氮配位,又同时与Tyr的氧配位形成氧桥。漆酶的底物专一性不强,据1989年初步统计漆酶的底物已超过200个。按底物结构可分为5类:(1)酚类及其衍生物,它们约占漆酶底物总数一半以上,包括邻苯二酚、对苯二酚、间苯二酚、α及β萘酚、硫酚、甲基酚及其衍生物等,真菌漆酶还可催化一元卤代酚的氧化。一元酚衍生物取代基一般在酚羟基的邻、对位。随取代基数目、种类和位置不同,反应活性也有差异。如漆酶催化下列一元取代酚氧化的活性次序分别为:P-甲酚>0-甲酚>m-甲酚》苯酚;2,6-二甲氧基酚>2,6-二甲基酚>邻甲氧基酚>邻甲基酚。(2)芳胺及其衍生物,它们的结构除氨基外与酚类相似,还有N,N-二甲基对苯二胺,1,5-萘二胺,联苯胺等。(3)羧酸及其衍生物,局限于芳环连有羟基或氨基的芳香酸、碳链连有酚或芳胺基团的羧酸及其酯。如原儿茶酸、原儿茶酸甲、丁、戊酯,3,4-二羟基苯乙酸等。(4)甾体激素和生物色素,在有机溶剂中真菌漆酶可催化氧化雌甾二醇、α-卵胞激素等激素,漆树酶可催化氧化胆红素、苏木色精、茜素红等色素。(5)其他非酚类底物:包括维生素C,亚铁氰化钾、二茂铁、二茂铁衍生物(Fc-R,R=CH2OH,CH2NEt2,COO-,COOCH3,CH2Fc等),以及1,2,4,5-四甲氧基苯和1-苯基-2-(3,4-二甲氧基)乙二醇等。漆酶催化氧化反应专一性较差,它催化酚(或芳胺)氧化时,酚羟基或氨(胺)基上先失去一个电子生成自由基,后者再发生一系列非酶反应,氧化成醌,发生键的断裂,导致聚合或偶联反应。1960年,中村隆雄用ESR监测对苯二酚的酶促氧化,发现反应分为两步:(1)底物酚向漆酶转移一个电子生成半醌-氧自由基中间体,在ESR谱上出现明显的吸收信号。(2)不均衡的非酶反应,二分子半醌生成一分子对苯醌和一分子氢醌。半醌氧自由基可以转变为碳自由基,继而发生碳碳或碳氧偶联生成较复杂的产物。在漆酶催化氧化过程中铜起决定性作用。但伯查德却认为是锰起决定作用。1986年,莱恩(Lehn)等用X-射线荧光光谱分析技术对伯查德使用过的漆树酶样品和他们制备的纯漆酶样品进行金属含量分析比较,发现伯查德的漆酶样品中杂质锰含量较高,因而导致错误的结论。漆酶是含铜单电子氧化还原酶,其中Ⅰ型铜和Ⅱ型铜是单电子受体,Ⅱ型铜是双电子受体,催化底物氧化时底物首先将电子传递给Ⅰ型铜和Ⅱ型铜,然后通过分子内的电子转移给Ⅲ型铜,在Ⅲ型铜上氧被还原为水。这样通过3种类型4个铜离子的协同作用,传递电子,同时发生铜离子的价态变化,实现漆酶对底物的催化氧化。对漆树酶中哪一类铜首先从底物接受电子存在分歧,美国哈尔德(Holwerda)和格雷(Gray)认为是Ⅱ型铜首先从底物接受电子,而瑞典恩决森(Andresson)和瑞哈姆(Reinhammar)则认为是Ⅰ型铜,产生分歧的原因是对实验现象解释不同。但他们对真菌漆酶催化氧化反应中的电子转移认识一致,都认为是Ⅰ型铜从底物酚中首先接受电子。真菌漆酶的活性比漆树酶大,因为后者的Ⅰ型铜被深埋在酶蛋白的多肽链内而显示出较小的反应活性。漆酶的用途除催化漆酚氧化使生漆干燥成膜外,还可作毛发染色的催化剂、助染剂、果实催熟剂、啤酒和果汁的澄清剂、分析和有机合成试剂等,都是基于漆酶的催化氧化反应。例如染色剂中的酚或芳胺经漆酶催化在头发上氧化成醌类物质,使头发呈橙黄色,染色程度可以通过加入的酶和染色剂量来控制。70年代以后固定化漆酶的制备为漆酶的实际应用开拓了广阔的前景。1986年,鲍拉格(Bollag J.M.)等用固定化漆酶处理纸厂废水,可以有效地除去甲基酚等毒物。用琼脂等固定的漆酶电极测定多酚、多芳胺浓度达2×10-5~7×10-4mol/L;用戊二醛交联的固定化漆酶制成膜型或网状电极测定脑神经组织儿茶胺,能消除维生素C的干扰,具有较高的灵敏度、重复性和稳定性。100多年来,漆酶的研究久盛不衰,涉及化学、生物学、物理学、医学、环境等多种学科。尤其是20世纪70年代以后,ESR、NMR、IR、UV、MS、HPLC等现代科学技术的普遍应用,使漆酶的研究取得很大进展。改进了纯化方法得到高纯度的漆酶;漆酶的活性由定性检测发展到定量快速测定;初步弄清漆酶的化学组成和铜的作用;制备了脱辅基(T2D)漆酶,探索了漆酶的部分取代、再生和重建。尤其是制备了固定化漆酶,初步开拓了它的实际应用途径。但至今没有得到漆酶的单晶,它的三维空间催化活性中心的立体结构还不清楚;漆酶催化反应机理和它在木质素生物合成或降解中的作用还缺乏直接、充分有力的证据;漆酶作为一个氧化还原酶还没有为人类充分地利用,有待于人们做深入细致的艰苦研究,才能取得更大的进展。【参考文献】:1 Reinhammar B,Laccase. in Copper Proteins and Copper En-zymes. (Lontie R,Ed)Vol II , CRC Press Boca Raton,1984: 1~ 352 Felici M,et al. J Chromatogr, 1985,320:4353 Mayer A M. Pvtchemistry,1987,26(1) :113 MayerAM.Pytchemistry,1987,26(1)∶114 季立才.中国生漆,1990,4∶225 Meadows K A,et al.J Inorg Biochem,1991,41(4)∶253~606 Galliano H,et al.Enzyme Microb Technol,1991,13(6)∶478(武汉大学季立才副教授撰) |