【果树原生质体研究进展】
拼译:advances in protoplast of fruit crops
柑桔是果树中原生质体培养研究最早、进展最快的树种。1975年,以色列的Vardi等以Shamouti甜橙珠心组织诱导胚性愈伤组织,并以此为起始材料分离原生质体,经培养,通过胚状体途径再生出植株。之后,他们又获得了甜橙、椪柑、柠檬等多个柑桔品种的原生质体再生植株。随后,日本的Kobayashi等(1983)也由Trovita甜橙分离出原生质体,并得到了再生植株。我国邓秀新等(1988、1989)从锦橙和山金柑珠心诱导的愈伤组织分离原生质体,并培养再生出植株。迄今,柑桔原生质体培养技术已被世界上许多实验室所掌握,并成功地应用于进一步的细胞融合、突变体筛选和遗传转化的研究中。柑桔原生质体分离与培养技术的建立和完善,为原生质体操作在柑桔育种上的应用创造了条件,也为其它木本果树的原生质体培养研究积累了经验。落叶果树的原生质体培养最先是在梨属植物上获得成功的。1986年,Ochatt等报道了野梨叶肉原生质体再生植株。不久,在樱桃、猕猴桃、苹果等果树上也获得了成功。80年代后期以来,果树原生质体培养取得了突破性进展,成功的树种不断增多。到目前,人们已经获得了包括柑桔、苹果、梨、葡萄、猕猴桃等大宗果树在内的9科15属果树的再生植株(表1)。桃、银杏也获得了愈伤组织或胚状体。
表1 果树原生质体培养获得再生植株概况


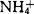 (达20.6mmol/l)。研究表明,高浓度的
(达20.6mmol/l)。研究表明,高浓度的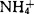 对原生质体有毒害作用,梨属植物表现尤为敏感。在含
对原生质体有毒害作用,梨属植物表现尤为敏感。在含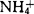 的MS培养基中,Bartlett梨原生质体仅能再生细胞壁;去掉其中的
的MS培养基中,Bartlett梨原生质体仅能再生细胞壁;去掉其中的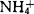 ,细胞虽能分裂,但到10个细胞时就死亡;只有在去掉
,细胞虽能分裂,但到10个细胞时就死亡;只有在去掉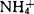 ,添加50mg/l水解酪蛋白(CH)的培养基中,原生质体才能持续分裂。在野梨、Conference、Passe Crassane、Old Home等梨品种上也得到类似的结果。因此很多实验室降低或完全去掉培养基中的
,添加50mg/l水解酪蛋白(CH)的培养基中,原生质体才能持续分裂。在野梨、Conference、Passe Crassane、Old Home等梨品种上也得到类似的结果。因此很多实验室降低或完全去掉培养基中的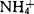 。Grosser等人(1987)取消了MT基本培养基本培养基的无机态氮,采用谷氨酰胺为氮源,建立了BH3培养基,大大提高了柑桔原生质体的分裂频率,已成为柑桔原生质体培养中较为通用的培养基。原生质体生长与发育需要多种维生系和有机成分。然而,并非所有果树都需要复杂的有机附加物。Conferene梨原生质体在加有KM8P有机物的培养基中分裂数次后便停止生长,樱桃上也看到类似的现象。众多研究表明,复杂的有机成分利于苹果和梨砧木原生质体培养。(chatt(1991)在培养基中添加甘氨酸(0.1~0.3mol/1),或与PVP(0.5%)混用,有效地防止了甜樱桃再生细胞团的褐化。同组织和细胞培养一样,外源生长调节剂对果树原生质体培养也是必需的,一般包括生长素类和细胞分裂素类。各种果树对这两大类激素的种类和浓度的需求有所不同,但一致的情况是,培养前期保持较高水平的生长素/分裂素,才能启动细胞壁的再生和细胞分裂。作为一个例外,在不加任何外源激素的情况下,柑桔原生质体就能分裂,形成多细胞团,进而发育成胚状体。添加BA和NAA反而抑制了细胞团的形成,这与其它果树形成鲜明的对比。原生质体的接种密度对培养效果影响很大。随着密度提高,锦橙原生质体恢复分裂的比例明显增加。樱桃、苹果也是如此。但密度过高,再生细胞团很小,而且很快停止生长。一般认为,密度过高造成营养不良;过低,又由于细胞内含物的外渗而影响细胞生长。柑桔胚性愈伤组织原生质体以(0.8~1)×105个/ml为宜,草莓叶肉原生质体4×105个/ml,而樱桃叶肉原生质体要求达到5×105个/ml。Ochatt等(1988)用电脉冲处理Colt樱桃悬浮细胞原生质体,促进了细胞分裂和植株分化,再生植株的生根率也明显高于对照。他们在进行野梨与樱桃体细胞杂交试验时发现,融合前,亲本原生质体先经电脉冲处理后,融合体能持续分裂,并分化植株,而未经处理的,培养40d后就停止分裂。电脉冲处理,可能促进了细胞吸收外源激素和营养物质,刺激了DNA的合成。4.植株的再生。果树原生质体培养以2种方式再生植株:一是由胚状体发育成植株;二是从愈伤组织诱导形态分化。柑桔原生质体培养到肉眼可见的细胞团后,转入固体培养基上2~3周,即可形成胚状体。如果在摇床上培养(120r/min),可促进胚状体发生。培养基中加入硫酸腺嘌呤5mg/l或麦芽浸出物(ME)500mg/l,有助于胚状体的进一步发育。当胚状体长到一定大小时,转到MT附加2.0mg/1 GA3固体培养基上,胚状体可直接萌发成植株,但频率较低。在MT附加2.0~5.0mg/l BA的固体培养基上,产生丛生芽,通过试管内嫁接或自根苗方式形成完整植株。龙眼、香蕉和番木瓜也通过这种途径再生出植株。多数果树原生质体先发育成愈伤组织,然后分别诱导芽和根。在这一途径中,不定芽的诱导是关键步骤,主要是通过调节培养基中激素的种类与浓度来实现这一目的的。愈伤组织发育到0.15~0.5cm大小时,转入固体分化培养基上诱导器官形成。分化培养基不加高浓度的渗透剂,只需一定浓度的糖作碳源,通常为2%一3%蔗糖,但枇杷只在3%或5%山梨糖醇的培养基上才能分化茎芽。提高分裂素/生长素水平利于芽的分化。苹果、梨原生本再生的愈伤组织在MS(或1/2MS)附加1.0~2.0mg/lBA和0.05~0.1mg/l NAA或1AA上,可产生芽,而樱桃需要玉米素、BA和NAA的配合使用。TDZ在低浓度时具有细胞分裂素的功能,对苹果、草莓芽的分化有促进作用。猕猴桃原生质体植株分化是分步进行的。经过几次过渡培养,逐步提高玉米素的浓度,降低2,4-D的浓度,最终去掉2,4-D。这样,给愈伤组织提供一个适应的过程,以利于器官分化。但张远记等(1995)将毛花猕猴桃愈伤组织直接从原生质体培养基中转到固体分化培养基上,也成功地得到了植物。通过外源激素的调控,愈伤组织也可先分化根,然后再生茎芽。欧洲酸樱桃CAB4D叶肉原生质体再生的愈伤组织。在MS附加0.025mg/l NAA,1.0mg/l BA和0.1mg/l ZT上发出根。切去根尖,将根段接入MS附加0.01mg/l NAA,2.0mg/l BA中,从根上长出植株。虽然果树原生质体培养已经取得了长足的进展。但是,除柑桔外,在其它果树上,再生植株的例证尚不够多,即使成功,也多为实生或砧木类型,而生产上大量推广的优良品种,很少见到原生质体培养再生植株的报道。另外,一些重要的果树,如桃、东方梨、梅、枣等也未成功。在获得再生植株的果树上,原生质体的来源也极为有限。柑桔等仅限于以胚性愈伤组织或胚性细胞系为材料,而苹果、梨、草莓等也只得到了叶内、幼茎或叶柄来源的原生质体的再生植株。今后,在总结已有经验的基础上,应扩大供试材料的类型和基因型的范围,着重研究各种来源的原生质体分离与培养条件,争取更多的树种、品种获得成功,为大规模地应用原生质体操作技术进行果树品种的改良奠定基础。原生质体融合 在果树上,柑桔原生质体融合研究最为领先。1985年,日本Ohgawara等采用PEG(聚乙二醇)诱导融合,首次获得了柑桔体细胞杂种植株,这一结果标志着果树原生质体操作研究进入了新阶段。之后,成功的组合迅速增多,已有几十个组合再生出杂种植株,其中包括一些亲缘关系较远的组合,如黄皮与柑桔,非洲樱桃桔与柑桔等。一批体细胞杂种已陆续开花。落叶果树原生质体融合相对滞后,报道也不多,到目前,仅有1例成功的试验。Ochatt等(1989)将野梨叶肉原生质体与樱桃悬浮细胞原生质体融合,获得了亚科间体细胞杂种。其他果树,如苹果、草莓、番木瓜,也进行了原生质体融合的探索,但尚未有再生植株的报道。该技术采用甲亲本的愈伤组织或悬浮细胞原生质体与乙亲本叶肉原生质体为材料,经PEG诱导融合。近来,Hidaka等(1992)、Shinozaki等(1992)利用电场诱导融合,得到几个柑桔体细胞杂种。融合后的原生质体,经再生技术体系培养成植株。在培养过程中,由于没有一个十分有效的杂种选择体系,再生植株必须经过鉴定才能确定是否为体细胞杂种。目前主要采用植株形态、染色数目、同工酶谱和DNA图谱等指标进行鉴定。体细胞杂交后,把所有遗传物质都融合在一起,很难排除不利性状的进入。胞质杂种巧妙地解决了这一难题。Vardi等(1987)应用供/受体融合系统,得到了具有枳橙胞质和酸橙细胞核的胞质杂种。华中农业大学柑桔研究所也正在进行这方面的研究工作。三倍体在果树生产上具有特殊的应用价值。苹果、梨、香蕉等果树的主栽品种中,三倍体品种占有重要的地位。除了传统育种方法外,应用原生质体融合及其下游技术也可培育出三倍体品种。目前,主要通过二种途径实现这一目标:一是用一亲本的体细胞原生质体与另一亲本的性细胞(如四分体小孢子)原生质体融合,培养再生,一步获得三倍体类型。与有性杂交相比,省去了培养四倍体的时间,因而大大缩短了育种周期。通常把综合性状优良的亲本作为二倍体一方,而具有个别特殊性状的亲本为单倍体。二是将体细胞杂种与二倍体品种杂交,也可培育出三倍体品种。Oiyama等(1991)将Trovita甜橙与枳的体细胞杂种与克里迈丁桔杂交,得到了三倍体植株。华中农业大学柑桔研究所运用这一途径,也获得了三倍体植株。无性系变异筛选、离体诱变及遗传转化原生质体呈单细胞状态,每个植株都是单细胞起源的,每个原生质体都有同等的再生机会。因而,原生质体培养为突变体的分离,纯化及离体诱变提供了便利条件。广东省果树研究所与华中农业大学合作,正在利用这一技术纯化红江橙红肉型自然突变嵌合体。美味猕猴桃愈伤组织原生质体再生植株在叶片、节间及花的形态上表现出大量的无性变异,其中还出现了矮生变异。更为有趣的是,从同一雌株诱导的愈伤组织分离出的原生质体,再生植株出现了雌雄性别分化。邓占鳖等(1989)应用原生质体培养体系,并结合人工诱变和离体筛选,获得了耐NaCl0.6%的‘桃叶橙’细胞系,并再生了植株,目前正进行田间观察。Gentile等(1992)利用原生质体再生体系,获得了抗黑星病(malsecco)的柠檬突变体。Kobayashi等(1989)用PEG诱导将带有氨基糖苷磷酸转移酶基因〔APH(3′)Ⅱ〕的细菌质粒DNA直接导入Trovita甜橙原生质体。用相同的方法,Oliveira等(1991)将带有氯霉素乙酰转移酶(CAT)标记基因的质粒DNA导入美味猕猴桃原生质体,也获得了瞬时表达。Nyman等(1992)采用电激法,将带有β-葡萄苷酸酶(GUS)标记基因的质粒DNA导入草莓叶肉和叶柄原生质体,并再生出了转基因植株。虽然遗传转化研究在果树上刚刚起步,而且所转化的多为标记基因,但它已显示出在果树育种上的应用潜力。可以预见,利用原生质体进行遗传转化研究,将对果树品种改良起到积极的推动作用。【参考文献】:1 邓秀新,章文才.柑桔四分体小孢子原生质体的分离.中国柑桔.1990,(1):7~8
。Grosser等人(1987)取消了MT基本培养基本培养基的无机态氮,采用谷氨酰胺为氮源,建立了BH3培养基,大大提高了柑桔原生质体的分裂频率,已成为柑桔原生质体培养中较为通用的培养基。原生质体生长与发育需要多种维生系和有机成分。然而,并非所有果树都需要复杂的有机附加物。Conferene梨原生质体在加有KM8P有机物的培养基中分裂数次后便停止生长,樱桃上也看到类似的现象。众多研究表明,复杂的有机成分利于苹果和梨砧木原生质体培养。(chatt(1991)在培养基中添加甘氨酸(0.1~0.3mol/1),或与PVP(0.5%)混用,有效地防止了甜樱桃再生细胞团的褐化。同组织和细胞培养一样,外源生长调节剂对果树原生质体培养也是必需的,一般包括生长素类和细胞分裂素类。各种果树对这两大类激素的种类和浓度的需求有所不同,但一致的情况是,培养前期保持较高水平的生长素/分裂素,才能启动细胞壁的再生和细胞分裂。作为一个例外,在不加任何外源激素的情况下,柑桔原生质体就能分裂,形成多细胞团,进而发育成胚状体。添加BA和NAA反而抑制了细胞团的形成,这与其它果树形成鲜明的对比。原生质体的接种密度对培养效果影响很大。随着密度提高,锦橙原生质体恢复分裂的比例明显增加。樱桃、苹果也是如此。但密度过高,再生细胞团很小,而且很快停止生长。一般认为,密度过高造成营养不良;过低,又由于细胞内含物的外渗而影响细胞生长。柑桔胚性愈伤组织原生质体以(0.8~1)×105个/ml为宜,草莓叶肉原生质体4×105个/ml,而樱桃叶肉原生质体要求达到5×105个/ml。Ochatt等(1988)用电脉冲处理Colt樱桃悬浮细胞原生质体,促进了细胞分裂和植株分化,再生植株的生根率也明显高于对照。他们在进行野梨与樱桃体细胞杂交试验时发现,融合前,亲本原生质体先经电脉冲处理后,融合体能持续分裂,并分化植株,而未经处理的,培养40d后就停止分裂。电脉冲处理,可能促进了细胞吸收外源激素和营养物质,刺激了DNA的合成。4.植株的再生。果树原生质体培养以2种方式再生植株:一是由胚状体发育成植株;二是从愈伤组织诱导形态分化。柑桔原生质体培养到肉眼可见的细胞团后,转入固体培养基上2~3周,即可形成胚状体。如果在摇床上培养(120r/min),可促进胚状体发生。培养基中加入硫酸腺嘌呤5mg/l或麦芽浸出物(ME)500mg/l,有助于胚状体的进一步发育。当胚状体长到一定大小时,转到MT附加2.0mg/1 GA3固体培养基上,胚状体可直接萌发成植株,但频率较低。在MT附加2.0~5.0mg/l BA的固体培养基上,产生丛生芽,通过试管内嫁接或自根苗方式形成完整植株。龙眼、香蕉和番木瓜也通过这种途径再生出植株。多数果树原生质体先发育成愈伤组织,然后分别诱导芽和根。在这一途径中,不定芽的诱导是关键步骤,主要是通过调节培养基中激素的种类与浓度来实现这一目的的。愈伤组织发育到0.15~0.5cm大小时,转入固体分化培养基上诱导器官形成。分化培养基不加高浓度的渗透剂,只需一定浓度的糖作碳源,通常为2%一3%蔗糖,但枇杷只在3%或5%山梨糖醇的培养基上才能分化茎芽。提高分裂素/生长素水平利于芽的分化。苹果、梨原生本再生的愈伤组织在MS(或1/2MS)附加1.0~2.0mg/lBA和0.05~0.1mg/l NAA或1AA上,可产生芽,而樱桃需要玉米素、BA和NAA的配合使用。TDZ在低浓度时具有细胞分裂素的功能,对苹果、草莓芽的分化有促进作用。猕猴桃原生质体植株分化是分步进行的。经过几次过渡培养,逐步提高玉米素的浓度,降低2,4-D的浓度,最终去掉2,4-D。这样,给愈伤组织提供一个适应的过程,以利于器官分化。但张远记等(1995)将毛花猕猴桃愈伤组织直接从原生质体培养基中转到固体分化培养基上,也成功地得到了植物。通过外源激素的调控,愈伤组织也可先分化根,然后再生茎芽。欧洲酸樱桃CAB4D叶肉原生质体再生的愈伤组织。在MS附加0.025mg/l NAA,1.0mg/l BA和0.1mg/l ZT上发出根。切去根尖,将根段接入MS附加0.01mg/l NAA,2.0mg/l BA中,从根上长出植株。虽然果树原生质体培养已经取得了长足的进展。但是,除柑桔外,在其它果树上,再生植株的例证尚不够多,即使成功,也多为实生或砧木类型,而生产上大量推广的优良品种,很少见到原生质体培养再生植株的报道。另外,一些重要的果树,如桃、东方梨、梅、枣等也未成功。在获得再生植株的果树上,原生质体的来源也极为有限。柑桔等仅限于以胚性愈伤组织或胚性细胞系为材料,而苹果、梨、草莓等也只得到了叶内、幼茎或叶柄来源的原生质体的再生植株。今后,在总结已有经验的基础上,应扩大供试材料的类型和基因型的范围,着重研究各种来源的原生质体分离与培养条件,争取更多的树种、品种获得成功,为大规模地应用原生质体操作技术进行果树品种的改良奠定基础。原生质体融合 在果树上,柑桔原生质体融合研究最为领先。1985年,日本Ohgawara等采用PEG(聚乙二醇)诱导融合,首次获得了柑桔体细胞杂种植株,这一结果标志着果树原生质体操作研究进入了新阶段。之后,成功的组合迅速增多,已有几十个组合再生出杂种植株,其中包括一些亲缘关系较远的组合,如黄皮与柑桔,非洲樱桃桔与柑桔等。一批体细胞杂种已陆续开花。落叶果树原生质体融合相对滞后,报道也不多,到目前,仅有1例成功的试验。Ochatt等(1989)将野梨叶肉原生质体与樱桃悬浮细胞原生质体融合,获得了亚科间体细胞杂种。其他果树,如苹果、草莓、番木瓜,也进行了原生质体融合的探索,但尚未有再生植株的报道。该技术采用甲亲本的愈伤组织或悬浮细胞原生质体与乙亲本叶肉原生质体为材料,经PEG诱导融合。近来,Hidaka等(1992)、Shinozaki等(1992)利用电场诱导融合,得到几个柑桔体细胞杂种。融合后的原生质体,经再生技术体系培养成植株。在培养过程中,由于没有一个十分有效的杂种选择体系,再生植株必须经过鉴定才能确定是否为体细胞杂种。目前主要采用植株形态、染色数目、同工酶谱和DNA图谱等指标进行鉴定。体细胞杂交后,把所有遗传物质都融合在一起,很难排除不利性状的进入。胞质杂种巧妙地解决了这一难题。Vardi等(1987)应用供/受体融合系统,得到了具有枳橙胞质和酸橙细胞核的胞质杂种。华中农业大学柑桔研究所也正在进行这方面的研究工作。三倍体在果树生产上具有特殊的应用价值。苹果、梨、香蕉等果树的主栽品种中,三倍体品种占有重要的地位。除了传统育种方法外,应用原生质体融合及其下游技术也可培育出三倍体品种。目前,主要通过二种途径实现这一目标:一是用一亲本的体细胞原生质体与另一亲本的性细胞(如四分体小孢子)原生质体融合,培养再生,一步获得三倍体类型。与有性杂交相比,省去了培养四倍体的时间,因而大大缩短了育种周期。通常把综合性状优良的亲本作为二倍体一方,而具有个别特殊性状的亲本为单倍体。二是将体细胞杂种与二倍体品种杂交,也可培育出三倍体品种。Oiyama等(1991)将Trovita甜橙与枳的体细胞杂种与克里迈丁桔杂交,得到了三倍体植株。华中农业大学柑桔研究所运用这一途径,也获得了三倍体植株。无性系变异筛选、离体诱变及遗传转化原生质体呈单细胞状态,每个植株都是单细胞起源的,每个原生质体都有同等的再生机会。因而,原生质体培养为突变体的分离,纯化及离体诱变提供了便利条件。广东省果树研究所与华中农业大学合作,正在利用这一技术纯化红江橙红肉型自然突变嵌合体。美味猕猴桃愈伤组织原生质体再生植株在叶片、节间及花的形态上表现出大量的无性变异,其中还出现了矮生变异。更为有趣的是,从同一雌株诱导的愈伤组织分离出的原生质体,再生植株出现了雌雄性别分化。邓占鳖等(1989)应用原生质体培养体系,并结合人工诱变和离体筛选,获得了耐NaCl0.6%的‘桃叶橙’细胞系,并再生了植株,目前正进行田间观察。Gentile等(1992)利用原生质体再生体系,获得了抗黑星病(malsecco)的柠檬突变体。Kobayashi等(1989)用PEG诱导将带有氨基糖苷磷酸转移酶基因〔APH(3′)Ⅱ〕的细菌质粒DNA直接导入Trovita甜橙原生质体。用相同的方法,Oliveira等(1991)将带有氯霉素乙酰转移酶(CAT)标记基因的质粒DNA导入美味猕猴桃原生质体,也获得了瞬时表达。Nyman等(1992)采用电激法,将带有β-葡萄苷酸酶(GUS)标记基因的质粒DNA导入草莓叶肉和叶柄原生质体,并再生出了转基因植株。虽然遗传转化研究在果树上刚刚起步,而且所转化的多为标记基因,但它已显示出在果树育种上的应用潜力。可以预见,利用原生质体进行遗传转化研究,将对果树品种改良起到积极的推动作用。【参考文献】:1 邓秀新,章文才.柑桔四分体小孢子原生质体的分离.中国柑桔.1990,(1):7~8(华中农业大学作物遗传改良国家重点实验室史永忠、邓秀新撰)